Синтез и распад белка в скелетных мышцах человека во время и после упражнений

Авторы: Vinod Kumar, Philip Atherton, Kenneth Smith, Michael J. Rennie.
Перевод — Сергей Струков.
Скелетные мышцы демонстрируют удивительную пластичность в ответ на изменения типа, продолжительности и интенсивности нагрузки, которые могут вызвать гипертрофию или атрофию, ограниченную гиперплазию и дифференциальную экспрессию множества белков и даже целых органелл, таких как митохондрии, с результирующими изменениями метаболизма субстратов и белков. С целью лучшего описания упражнения традиционно подразделяются на «аэробные / на выносливость» и «с отягощением (силовые)». Основное видимое отличие упражнений на выносливость (то есть повторяющихся низкоинтенсивных сокращений, которые могут выполняться продолжительное время) – смещение фенотипа в сторону возрастания количества волокон с более высокими окислительными способностями, в то время как повторные упражнения с отягощением (представляющие собой сокращения высокой интенсивности) вызывают гипертрофию волокон (и, вероятно, некоторую гиперплазию вследствие активации клеток-сателлитов). В реальности присутствует существенное «перекрытие» между вариантами ответов, это происходит вследствие восприятия и распределения мышцей сигналов в ответ на активность, которые приводят к постоянной адаптации к специфичной природе, интенсивности и продолжительности упражнений. В предлагаемом обзоре упражнения будут разделяться на «резистивные» (с дополнительным отягощением) и «нерезистивные» (без отягощения)…
…Целью обзора является описание регуляции обмена мышечных белков у человека во время и после упражнения, влияния на него факторов окружающей среды, таких как тип нутриентов, состав и уровень потребления, а также полового диморфизма и возраста. В обзоре представлены следующие темы:
- методики изучения обмена в мышцах человека;
- упражнения и общий мышечный белковый баланс;
- синтез мышечного протеина (СМП) и упражнения с отягощением (во время и после упражнения);
- СМП в упражнениях без отягощения (во время и после);
- распад мышечного протеина (РМП) и упражнения (во время и после упражнений с отягощениями и без них);
- влияние пищи (и синергия с упражнениями) на обмен мышечных белков;
- влияние продолжительной тренировки на метаболизм мышечных белков;
- пол и упражнения;
- старение и упражнения.
Мы включили в обзор три таблицы, в которых суммированы наиболее исчерпывающие исследования в данной области, проведенные в последние два-три десятилетия, освещающие специфичные переменные: возраст и пол, тип выполняемых упражнений, нутрициональные манипуляции, а также изменения в синтезе и распаде белков, позволяющие читателю ознакомиться с работами в данной области. Мы также включили рисунок, схематически отображающий основные клеточные сигнальные пути, вовлеченные в регуляцию синтеза мышечного белка в ответ на упражнения и прием пищи, – так, где возможно, мы кратко опишем связанные с этим изменения активности регуляторных путей как итог изменений их статуса фосфорилирования.
Мы ограничимся явлениями, происходящими в течение минут и часов (реже дней, исключительно для эффектов тренировки) во время или после упражнения. Изменения во время упражнения отражают активацию метаболических приоритетов в направлении переноса энергии для мышечной работы при множественных изменениях, таких как ингибирование синтеза белка, увеличение трансаминирования и окисления аминокислот – скорее сопровождающее явление, нежели специфическая функциональная адаптация в ответ на упражнения. Изменения в обмене мышечного белка в период после упражнений больше отражает приспособительное ремоделирование (например, увеличение синтеза группы миофибриллярных протеинов, ведущей к гипертрофии). Мы не будем обсуждать изменения в уровнях транскрипции генов.
Упражнения и общий баланс белка в мышцах
Мышечная масса поддерживается путем регулирования баланса между СМП и РМП. Общее увеличение мышечной массы возможно только в случае, если СМП превышает РМП, то есть при положительном белковом балансе, и наоборот. Во время отдыха в состоянии голода (правильнее называть постабсорбтивное состояние) общий баланс белка отрицательный, а позитивный баланс обычно наблюдается только, когда во время еды восполняются потери мышечного белка, поддерживая стабильную массу мышц. После упражнений в состоянии голода, независимо от увеличения СМП (см. ниже), общий баланс белка становится даже более негативным, не достигая положительных величин, так как уровень РМП, повышенный перед упражнениями, также повышается (13). Тем не менее, когда протеин или аминокислоты потребляются после тренировки, общий баланс белка становится положительным, так как СМП подавляет РМП, который может быть пониженным сам по себе (111).
СМП и упражнения с отягощениями
Во время упражнений

Измерения СМП человека, сделанные во время выполнения упражнений с отягощениями, редки, так как большинство исследований применяют тренировку короче минимального периода времени (около часа), которое требуется современным методикам для оценки проявления строгих изменений меченных стабильных изотопов. Также упражнения прерывистые, используются подходы повторяющихся сокращений, разделенные периодами отдыха, при которых мышца не пребывает в состоянии покоя, что усложняет интерпретацию полученных данных, особенно при методе, основанном на A-V пробах и кровотоке. Данные исследований мышц грызунов (24) и людей (37, 43) показывают угнетение СМП во время упражнений с отягощением. В то же время в работах, использовавших метод растворенных A-V меток, не увеличивался уровень поглощения меток, то есть синтез белка в мышцах ног не изменялся (39). Противоречие между ранними исследованиями (37) и более поздними данными (39), возможно, вызвано методологическими различиями (использование метода прямого включения против метода растворенных меток A-V) или является результатом различий в объеме выполненной работы. Обнаруженное резкое понижение СМП вызвано сокращением этапов инициации и элонгации при трансляции мРНК (63) из-за уменьшения фосфорилирования связывающего белка 4Е 1 (4ЕВР-1), эффектора, запускаемого мишенью рапамицина млекопитающих (mTOR), и тенденции к увеличению фосфорилирования фактора элонгации эукариотов 2 (eEF2) – отрицательного регулятора элонгации пептидной цепи (37) (рис. 1).

Рис. 1. МАРК – митогенактивированная протеинкиназа; АМРК – АМФ-активированная протеинкиназа; Akt – протеинкиназа В; mTORC1 – мишень рапамицина млекопитающих С1; eIF4E-BP1, eIF2Bε, eIF4F и eIF2: белок, связывающий фактор инициации эукариотов 4Е, 2Bε, 4Fb 2; p70S6K1 – 70-кДа S6 протеинкиназа 1; rps6 – рибосомный протеин s6; eEF2 – фактор элонгации эукариот 2; 5’TOP, 5-терминальный олигопиримидин; tRNAi – инициатор тРНК.
Bylund-Fellenius и коллеги (24) наблюдали вызванные сокращением понижение СМП в перфузированной электростимулированной мышце крысы, активность АТФ миозина вызвала повышение коэффициента АМФ/АТФ, вследствие чего, вероятно, оказывается стимулирующее воздействие на активность АМФ-активированной протеинкиназы (АМРК) (52), что вызывает ингибирование сигнальных эффектов комплекса туберозного склероза 2 на mTOR и уменьшает фосфорилирование 4ЕВР-1 (18). Активность АМРК действительно повышается на >30 % после выполнения упражнений с отягощением (37), но важность ингибирования mTOR при выполнении упражнений не определена, так, синтез белка повышается после выполнения упражнений независимо от продолжающегося повышения фосфорилирования АМРК (37).
Недавние работы также продемонстрировали эффективность предварительного потребления аминокислот на СМП во время упражнения. После ночного голодания испытуемые приняли 0,35 г/кг сухой массы тела незаменимых аминокислот и 0,5 г/кг сухой массы тела сахарозы за час до упражнений (10 х 10, 70 % РМ). СМП понизился во время выполнения упражнений, но не ниже исходного уровня, наблюдаемого в контрольной, не потреблявшей добавок группе (43). Подобным образом прием белка перед и во время 2-часовой прерывающейся тренировки с отягощениями, включающей упражнения для всего тела, повышал общий белковый баланс и СМП во время выполнения упражнений (7).
После упражнений
Общепринято, что упражнения с отягощениями вызывают увеличение СМП в восстановительном периоде (29, 71, 131). Действительно, тренировка с отягощением может увеличивать уровень СМП после упражнений в 2–5 раз, и этот эффект наблюдается более 48 часов у сытых субъектов (85). В других сообщениях (37, 100) обнаруживали угнетение СМП непосредственно после напряженной сократительной активности. В нашей лаборатории обычно наблюдаемые изменения не превышали исходные при оценке включения меток в белки первые 1–1,5 часа (66), тогда как другие этого не делали (37, 100), тем не менее большинство оценивающих исследователей свидетельствуют, что любое повышение после упражнения меньше вначале и увеличивается впоследствии (38, 66). Стимуляция синтеза белка у нетренированных людей после упражнений с отягощениями происходит в митохондриальном и миофибриллярном пулах (123). Более того, мы недавно исследовали влияние тренировки с отягощением на СМП в широком диапазоне интенсивности упражнений: от 20 до 90 % РМ при эквивалентной общей работе (1620–1800 единиц) в постабсорбтивном состоянии у молодых (24 ± 6 лет) и пожилых (70 ± 5 лет) мужчин в течение 4 часов после окончания занятия (66). Результаты показывают, что величина синтеза миофибриллярных белков зависит от интенсивности, увеличиваясь от 20 %, и достигает плато между 60 и 90 % РМ. Требуются дополнительные исследования для выяснения влияния объема работы и адаптивного ответа обмена мышечного белка при различной интенсивности нагрузки.
Молекулярный механизм стимулирующего эффекта упражнений с отягощением интенсивно изучался в последние годы – сначала на грызунах (2, 4, 19), а когда усовершенствовалась аналитическая техника – на человеческих мышцах (1, 34, 36, 37, 61, 66). Доступные в настоящее время данные, наиболее точно описывающие сигнальные пути во время и непосредственно после упражнений в состоянии голода и насыщения, представлены на рис. 1. Детальное описание активации процессов фосфорилирования или активности сигнальных молекул, регулирующих клеточный СМП в ответ на упражнения с отягощением, выходит за рамки настоящего обзора. Вкратце активация сигнальных молекул, регулирующих трансляцию и синтез белка, таких как Akt (протеинкиназа В), митогенактивируемая протеинкиназа, mTOR и его эффекторы, такие как эукариотический фактор инициации 4EBP-1, p70S6k1 (70-кДа протеинкиназаS6) и рибосомная протеинкиназа s6, связаны с увеличением СМП в период после упражнения (1, 36, 37. 61, 66). Тем не менее, необходимо выяснить временную зависимость и продолжительность этих ответов, а также дозозависимые характеристики, как и потенциальную роль в адаптивном ответе мышц на однократные и хронические упражнения.
В отношении клеточных сигнальных путей мы полагаем, что в настоящее время у нас нет точного понимания связи между изменениями в сигналах и СМП или РМП. В частности, непонятно, действует ли молекула как простой переключатель сигнального пути или как усилитель, модулирующий результирующую метаболическую реакцию. Данные исследований действия инсулина на мышцы человека слишком упрощенны, чтобы объяснить отдельные физиологические влияния на синтез или распад белка путем воздействия на степень фосфорилирования рассматриваемых молекул (50). Эту информацию можно более элегантно отобразить на основе данных зависимости от дозы и времени, собранных в нашей лаборатории (16, 17, 66).
СМП и упражнения без отягощения
Во время упражнений

Вероятно, во время бега СМП у грызунов понижается (35). Более поздние исследования зафиксировали уменьшение СМП на 26 % у крыс во время 2-часового бега на тредмилле (46). В основе изменений СМП во время 30-минутного бега мышей на тредмилле, возможно, лежит повышение активности АМРК и подавление общего уровня трансляции мРНК, а также сигнального пути mTOR (125). У людей общий уровень синтеза белка понижался во время ходьбы в гору при 40 % максимального потребления кислорода (VO2max) (93), и подобные изменения наблюдаются во время 2-часовой ходьбы при 60 % VO2max (22). СМП является значительной частью общего синтеза белка организмом, также известно, что отношение АДФ к АТФ значительно уменьшается при упражнениях без отягощения (24), на этом основании можно сделать вывод: во время ходьбы или бега у людей понижается СМП, но практически нет данных, подтверждающих это. Фактически во время ходьбы на тредмилле при 40 % VO2max (27) нет существенных изменений в СМП сравнительно с уровнем во время отдыха, тем не менее исходный уровень мог быть необычно низким по отношению ко дню без тренировки, что, вероятно, привело к ложно негативным выводам. Также, возможно, выбран недостаточный уровень интенсивности упражнений: велоэргометрия в течение часа при 70 % VO2max, выполняемая молодыми здоровыми субъектами, приводила к активации мышечной АМРК-α2 (42), измеренной в квадрицепсе с помощью биопсии непосредственно после упражнения; в подобном исследовании продолжительностью 90 минут отмечена значительная, вызванная Са2+ активация кальмодулинзависимой протеинкиназы еЕF2 киназы, совместно (и, возможно, как результат) с ингибированием активности еЕF2 и (в итоге) удлинение белковой цепи у здоровых мужчин в постабсорбтивном состоянии (97). Эти результаты противоречат гипотезе о снижении СМП во время езды на велосипеде и бега, тем не менее технически сложно организовать исследование, в котором у субъектов, упражняющихся продолжительное время и со значительной нагрузкой, можно удовлетворительно отследить обмен белка во время упражнения, но это возможно. Задача ожидает своего решения.
После упражнений
После ходьбы на тредмилле при 40 % VO2max в постабсорбтивном состоянии СМП в различных мышцах увеличивался на 45 % (26); о подобных изменениях сообщал и Sheffield-Mooreetal (101). Еще большее увеличение уровня синтеза миофибриллярной фракции могут вызывать более интенсивные упражнения у сытых молодых мужчин, 1 час маховых упражнений ногами ̴̴ 70 % РМ удвоили уровень синтеза миофибриллярного белка в квадрицепсе на 24 часа, синтез оставался повышенным еще более 72 часов (76).
Эти результаты можно было бы считать случайностью, так как обычно упражнения этого типа (которые скорее увеличивают биогенез митохондрий) не вызывают гипертрофию и, следовательно, не стимулируют синтез миофибриллярных белков, но накопившиеся данные развеивают сомнения. Описанные нами факты обнаружены у нетренированных субъектов, в этом случае и отягощения, и велоэргометрия стимулируют синтез белков и миофибрилл и митохондрий, возможно, в результате общего анаболического сигнала после упражнений, в то же время у тренированных не повышается синтез миофибрилл после велоэргометрии и не наблюдается синтез митохондриальных белков после однократной тренировки с отягощениями (123).
Степень изменений СМП в ответ на упражнения, вероятно, зависит от того, производится или нет существенная «ударная» сила, например, после интенсивного плавания СМП у здоровых людей существенно не изменяется (113). Тем не менее, важно отметить, что измерения проводились после длительной тренировки, выполненной натощак. Кроме того, испытуемые были хорошо тренированы, а долговременная тренировка увеличивает базальный уровень СМП и уменьшает СМП в ответ на однократное воздействие упражнений (86, 87, 102).
В настоящее время существует много описаний активирующих фосфорилирование сигналов, которые могут лежать в основе изменений СМП после однократного выполнения упражнений без отягощения, а также изменений СМП самого по себе (например, увеличение сигналов mTOR, уменьшение еEF2, митогенактивирующей протеинкиназы и т. д.) (1, 11, 74, 98). Количественно наблюдаемые изменения сходны с происходящими в ответ на упражнения с отягощениями, и действительно, различия в ответ на однократные упражнения, при которых мышцы ног работают в разных режимах, «силовом» и «на выносливость», незначительны у тех же нетренированных людей (123). Это означает, что любое значительное увеличение сократительной активности или возможности утилизации субстратов нетренированными мышцами ведет к сходному анаболическому ответу. Тем не менее после периода тренировки СМП становится более чувствительным к специфическому типу упражнений, вызывая синтез отдельных субклеточных белковых фракций (митохондриальной или миофибриллярной), впоследствии проявляясь в фенотипических изменениях, характерных для разных видов тренировки (123). Фенотипические изменения, вероятно, возникают в результате повторного выполнения каждого из видов упражнений с отягощениями или «динамических». В настоящее время существует ограниченное количество данных о влиянии тренировки на активацию фосфорилирования сигнальных белков, этого недостаточно для возможности предсказания влияния на обмен протеинов изменений фосфорилирования.
РМП и упражнения
Во время упражнений с отягощениями
Только метод, основанный на A-V растворенных меченых аминокислотах, обсуждаемый ранее, доступен в настоящее время, однако его точность в данном случае весьма сомнительна. Насколько нам известно, только в двух исследованиях отслеживали уровень растворения меток «во время» упражнения (в действительности – во время отдыха между подходами) в постабсорбтивном состоянии; в этих исследованиях наблюдаемый уровень фенилаланина, показателя распада белка, не повышался во время отдыха (39, 114). Возможно, это объясняется тем, что протеолиз в мышцах происходит преимущественно через АТФ-зависимую убиквитинпротеазную систему (3), отношение АДФ к АТФ увеличивается во время упражнений, что может быть причиной его (распада) подавления во время выполнения упражнений, как в случае синтеза белка (см. выше).
После упражнений с отягощениями
Какой бы ни была неопределенность для периода упражнений, несомненно, что в постабсорбтивном состоянии после упражнения протеолиз в мышцах увеличивается, как показано с помощью растворенных меток (13, 15) и методом оценки уровня фракционного распада (ФРУ) (65). Перед упражнениями мышцы имели отрицательный баланс аминокислот, и эта ситуация лишь немного улучшалась при выполнении интенсивных упражнений с отягощениями, потому что, в то время как СМП увеличивался вдвое, ФРУ, существенно больший, чем ФРУ в постабсорбтивном состоянии, также увеличивался на 30–50 % спустя 3 часа, таким образом поддерживался отрицательный баланс (13, 85, 86). Тем не менее повышение распада в мышцах меньше по времени, чем уровень фракционального синтеза (24 часа по сравнению с 48) (85).
РМП и упражнения без отягощения
Неясны изменения РМП во время упражнений без отягощения. Во множестве экспериментов с циклическими упражнениями отмечали увеличение высвобождения аминокислот из нижних конечностей (72, 73) и на основании этого делали вывод о повышении протеолиза. Тем не менее высвобождение аминокислот объясняется скорее ингибированием синтеза белка, что приводит к недостаточной компенсации распада, и в результате из аминокислотного пула высвобождаются свободные аминокислоты.
Не вызывает сомнений, что в постабсорбтивном состоянии после упражнений без отягощений (45 минут ходьбы на тредмилле при 40 % VO2max) протеолиз в скелетных мышцах повышается как у молодых, так и у пожилых мужчин. В нижних конечностях увеличение протеолиза наблюдается через 10 и исчезает через 60 минут, но только у молодых субъектов (101).
Противоположные результаты получены с помощью микродиализа: концентрация 3МеН в диализате не изменялась от 6 до 72 часов после маховых упражнений одной ногой в течение часа при ̴ 70 % РМ (55). Возможно, это объясняется тем, что протеолизу подвергались «немиофибриллярные» протеины, при этом 3МеН не образуется, но более вероятно, что этот метод вообще не подходит для оценки мышечного протеолиза (13, 85, 95).
Сигнальные пути и РМП
Сигнальные пути, контролирующие РМП и протеолиз в мышцах человека, не определены. Различные протеолитические пути (включая лизосомальный, активируемый кальцием и убиквитин-протеосомо-зависимую систему, каспазы и металлопротеазы, а также неспецифические ди- и трипептидазы) вовлечены в ремоделирование мышц вследствие упражнений, но долевое участие каждого из них непонятно.
Увеличение активности активируемых кальцием протеаз и металлопротеаз отмечено в мышцах крысы во время и после бега на тредмилле (9, 25). Существует ограниченное количество сообщений об измерениях острых изменений производительности или контроля путей мышечного протеолиза у людей. В мышцах грызунов обнаружены две мышечные убиквитинлигазы, F-box мышечной атрофии (MAFbx) и мышечный reallyinterestingnovelgene fingerprotein 1 (MuRF1), стимулирующие протеолиз (3). Исследования экспрессии протеолитических генов в мышцах молодых субъектов спустя 4 часа после упражнений с отягощением, особенно экспрессии генов убиквитинапротеосом, показали увеличение активности MARFbxи MuRF1мРНК, но без существенных изменений в forkheadbox 3AмРНК (91). Парадоксально, но эксперименты нашей группы обнаружили снижение активности MARFbxм РНК в первые 24 часа после упражнений с отягощениями, которое означало увеличение РМП в посттренировочном состоянии (65). Это может быть связано с объемом и видом упражнения, выполняемого в последнем исследовании, где испытуемые выполняли подъемы на ступеньку с дополнительным отягощением, составлявшим 25 % от их массы тела, до полного утомления, а также со временем измерения. С другой стороны, в нашей лаборатории ранее не обнаружено прямой зависимости между РМП и экспрессией MARFbx (50). В любом случае существует и активируется множество путей, тем не менее интересно представить, как интактные миофибриллярные белки могут «реконструироваться» или ремоделироваться таким образом, чтобы оставалось место для вновь созданных протеинов, которые, согласно нескольким исследованиям (101, 113), синтезируются в течение часов после стимуляции упражнением.
Влияние потребления пищи на СМП и упражнения
 Потребление смешанной пищи усиливает СМП вдвое (94), и этот эффект, судя по нашим данным, обусловлен преимущественно действием аминокислот (10, 106), особенно лейцина (106), без значительного вмешательства инсулина (16, 32). Аминокислоты увеличивают синтез миофибриллярных, саркоплазматических, а также митохондриальных белков в скелетных мышцах (17), вероятно, дозозависимым способом (16, 32).
Потребление смешанной пищи усиливает СМП вдвое (94), и этот эффект, судя по нашим данным, обусловлен преимущественно действием аминокислот (10, 106), особенно лейцина (106), без значительного вмешательства инсулина (16, 32). Аминокислоты увеличивают синтез миофибриллярных, саркоплазматических, а также митохондриальных белков в скелетных мышцах (17), вероятно, дозозависимым способом (16, 32).
Упражнение с отягощением и еда синергически повышают СМП и ведут к положительному белковому балансу после упражнения, большему, чем просто потребление пищи (14). Несколько групп отметили, что потребление аминокислот или протеина, совместно с углеводами или без них, после упражнений с отягощением (14, 21, 33, 64, 77, 79, 90, 108, 112) или без отягощений (76) дополнительно усиливали СМП. Например, увеличение СМП на 145 % по сравнению с исходным состоянием происходило, когда после упражнений потреблялись обогащенные лейцином незаменимые аминокислоты с углеводами, тогда как без действия нутриентов СМП увеличивался на 41 % (36). Усиливающий эффект потребления пищи после упражнений, видимо, вызван увеличением количества аминокислот, а не глюкозы в крови (20, 77).
Кривая дозозависимого увеличения СМП после упражнения и потребления белка (80) имеет сходную форму с наблюдаемой в покое (17) с небольшим сдвигом влево вверх под влиянием упражнений. Таким образом, работы показывают синергизм между едой и упражнением. Также не выявлено преимуществ при потреблении большого количества белка (>20 г, что на самом деле немного) с целью повышения накопления мышечного протеина, максимально эффективная доза составляет, вероятно, 15–20 г высококачественного белка: мясного, яичного или соевого.
Время потребления пищи
Нет единого мнения, что приводит к большему СМП: потребление аминокислот до или после упражнений. Так, сообщалось (114), что прием незаменимых аминокислот с углеводами непосредственно перед упражнениями с отягощениями вызвал большее поглощение аминокислот нижними конечностями, но эти результаты трудно проверить количественно, так как они получены не в состоянии покоя и увеличение потребления некорректно отражает размер физиологического повышения (в 20 раз), вероятно, имеет место артефакт – захват аминокислот из внутримышечного пула. Более того, недавно показано путем прямого измерения синтеза отдельных фракций у людей, что потребление пищи за 1 час до интенсивных упражнений с отягощениями не приводит к дополнительному повышению СМП в течение 2 часов после окончания нагрузки (43). Таким образом, в очередной раз показано, что метод растворенных меток предоставляет количественно и качественно различающиеся результаты по сравнению с «включением» меченых аминокислот.
В то время как остается невыясненным, когда лучше потреблять белок для максимального СМП в ответ на упражнения, существуют результаты непрямых измерений в отношении долговременной тренировки, таких как гипертрофия, показывающие, что у молодых и пожилых людей происходит большее увеличение мышечной силы и массы, если белок потребляется непосредственно после упражнений, чем через несколько часов (40, 53, 67).
Качество белка
Большой интерес представляет предположение, что белки различного «биологического качества» и усвояемости могут быть более или менее эффективны для обеспечения мышц аминокислотами после упражнений. Недавние работы Филлипса и коллег (53, 124) выявили преимущество сывороточного белка перед казеином, соевым протеином и белком цельного молока относительно общего включения в белки мышц. В то же время нет доказательств в пользу преимуществ молочного протеина в сравнении мясным, яичным или соевым (78).
Анаболические сигнальные пути
Молекулярный механизм, лежащий в основе увеличения стимулирующего СМП эффекта еды после упражнений, связан с большим повышением фосфорилирования mTOR, p70S6K1 и 4EBP-1, чем только при выполнении упражнений (рис. 1) (36, 61, 64).
Влияние упражнений и приема пищи на РМП
Аминокислоты сами по себе оказывают, по крайней мере небольшое (50), ингибирующее влияние на распад белка в нижних конечностях у человека, особенно в присутствии инсулина, но эффект меньший, чем наблюдающийся у животных. Подавление разрушения белка в мышцах предплечья человека происходит после введения смеси аминокислот с разветвленной боковой цепью (68, 69). Большинство физиологических эффектов аминокислот на РМП в покое, вероятно опосредовано повышением секреции инсулина. Тем не менее несколько исследователей сообщили, что увеличение биодоступности аминокислот после упражнений не ингибирует существенно РМП человека (14, 21, 70, 90, 112).
Влияние тренировки на метаболизм мышечного белка
Продолжительные упражнения с отягощениями увеличивают средний поперечник волокон и вызывают гипертрофию мышц. Тем не менее мы преимущественно игнорируем временную зависимость изменений и точный механизм происходящего, который должен включать активацию СМП и ремоделирования, а также разрушение «отработавших» белков и РМП. Несколько работ отметили, что тренировка с отягощением увеличивает базальный уровень СМП (5, 86). Также было показано, что даже кратковременная (2 недели) тренировка с отягощениями увеличивает СМП в покое, но данные сложно интерпретировать в связи с тем, что СМП измеряли кратковременно (между 3 и 18 часами) после последнего занятия, и увеличение СМП могло быть следствием срочного влияния упражнений, а не тренировки самой по себе (54, 129, 131). В подтверждение увеличения СМП в покое вследствие упражнений зафиксировано повышение фосфорилирования Akt-mTOR-p70S6k (123). Тем не менее есть противоположные данные, полученные в той же лаборатории, в отношении долговременной тренировки (109). Мы с коллегами оценивали срочный эффект упражнений (велоэргометрия) с отягощением и без него в разных нижних конечностях одних и тех же субъектов после 10 недель тренировки на синтез миофибриллярных и митохондриальных белков: в ноге, тренировавшейся с отягощением, отмечено повышение синтеза миофибриллярного белка в состоянии покоя, тогда как тренировка без отягощения увеличила только синтез митохондриального белка (123). Эти результаты указывают на то, что повторные специфические нагрузки увеличивают синтез отдельных белковых фракций не в результате кратковременной модуляции активности трансляции, а за счет активации специфических программ транскрипции генов (65, 91).
Также отмечалось ингибирование СМП после однократной тренировки в результате долговременных упражнений (86). Тем не менее в той же лаборатории недавно обнаружено ̴ 48 % увеличение СМП после одной тренировки в результате 12 недель долговременного выполнения занятий, включающих упражнения с отягощениями (62). Эти различия объясняются относительно низким стимулом в тренированном состоянии вследствие применения аналогичной абсолютной интенсивности перед и после тренировки в предыдущем исследовании (62). Однако мы с коллегами также зафиксировали понижение синтетического ответа (̴ 30 %) после однократного занятия в конечности, «тренировавшейся» с отягощением, при аналогичной относительной интенсивности (123).
Имеются свидетельства не только различий величины ответных изменений СМП, но и временные различия: долговременные упражнения с отягощениями вызывают более быстрое, но кратковременное повышение СМП, чем однократное занятие у нетренированных (109). Таким образом, тренировочный статус – важный параметр, когда оценивается ответ мышц на однократные упражнения.
Сообщалось об увеличении СМП в латеральной широкой мышце в покое на 22 % в результате 16 недель велоэргометрии без отягощения (45 минут при 80 % максимальной ЧСС, 3–4 дня в неделю) (102). Вероятно, умеренное повышение общего фракционного синтеза белка было результатом более высокого увеличения синтеза в митохондриальной и/или саркоплазматической фракции. Даже спустя 4 недели после программы ходьбы/бега при 65–85 % максимальной ЧСС отмечался умеренно повышенный общий фракционный синтез белка (~17 %); тем не менее, отмечается парадоксальное явление: увеличение общего фракционного синтеза белка (~40 %) приводит к более отрицательному белковому балансу (87).
Если упражнения без отягощения вызывают увеличение СМП, почему же тогда не происходит гипертрофии мышц? Увеличение СМП после динамических упражнений может быть, в частности, связано с повышением синтеза белков, которые ответственны за адаптационные изменения при этом виде упражнений, то есть за увеличение объема митохондрий, активность митохондриальных энзимов и синтез белка в митохондриях (48, 57). В поддержку этого Шорт с коллегами сообщили об увеличении синтеза белков, транспортирующих глюкозу, белков митохондрий, уровня митохондриальных энзимов и 22 % увеличении общего СМП в покое вследствие 16-недельной программы «аэробных» упражнений (102, 103). Недавно Wilkinson et al. (123) показали, что повторные динамические упражнения продолжительностью более 10 недель увеличивают только синтез митохондриальных белков и не оказывают влияния на синтез белков миофибрилл или на общее фосфорилирование Akt-mTOR-p70S6k у молодых здоровых людей. На уровне транскрипции «негипертрофические» упражнения (30 минут бега при 75 % VO2max) увеличивают относительное содержание мРНК и транскрипцию различных миогенных и метаболических генов (для миогенной дифференциации гексокиназы II и пируватдегидрогеназной киназы 4) после упражнения, которое достигает максимума через 4–8 часов и возвращается к исходному уровню через 24 часа (126). Кумулятивным эффектом этого преходящего увеличения вследствие повторной динамической тренировки является адаптация мышц, наблюдаемая при упражнениях без отягощения (31).
Адаптивные изменения к динамическим упражнениям проявляются в подавлении АМРК, внеклеточной, регулирующей сигналы киназы 1/2 и активности сигналов mTOR после 10 дней интенсивных велоэргометрических нагрузок: 45–60 мин при 75–90 % у здоровых людей (11). Увеличение экспрессии специфичных мышечных коактиваторов транскрипции, активирующего пролиферацию рецептор-ү коактиватора-1α пероксисом (PPAR-1α), также может быть связано с адаптивным ответом на динамические упражнения, приводящим к биогенезу митохондрий и увеличению окислительной способности (88, 89). Тем не менее физиологическая роль PPAR-1α в адаптивном ответе мышц на тренировку нуждается в подробном изучении.
Половые различия в метаболизме мышечных белков и упражнения
 К сожалению, мало что известно про влияние полового диморфизма на композицию тела, при этом мужчины имеют большую массу мышц, чем женщины. Хорошо известно анаболическое влияние тестостерона на мышцы (41, 58), и секреция тестостерона в пубертатном периоде, скорее всего, ответственна за увеличение массы мышц в раннем взрослом возрасте. Тестостерон также увеличивает СМП в покое как у молодых, так и у пожилых людей (23, 116), но это влияние не связано со срочными изменениями синтеза белка, а проявляется ген-зависимыми изменениями, через воздействие на андрогенные рецепторы ядра. Женские половые гормоны ингибируют СМП и рост мышц у крыс (115), но не выявлено измеримых различий между мужчинами и женщинами в базальном уровне фракционного белкового синтеза или ответе на внутривенное введение аминокислот при умеренном уровне инсулина (Smith G., Mittendorfer B., Atherton P. и Rennie M. J., неопубликованные данные). Также не выявлены различия СМП или РМП в исходном уровне или после упражнений между молодыми мужчинами и женщинами (44, 59, 75, 82).
К сожалению, мало что известно про влияние полового диморфизма на композицию тела, при этом мужчины имеют большую массу мышц, чем женщины. Хорошо известно анаболическое влияние тестостерона на мышцы (41, 58), и секреция тестостерона в пубертатном периоде, скорее всего, ответственна за увеличение массы мышц в раннем взрослом возрасте. Тестостерон также увеличивает СМП в покое как у молодых, так и у пожилых людей (23, 116), но это влияние не связано со срочными изменениями синтеза белка, а проявляется ген-зависимыми изменениями, через воздействие на андрогенные рецепторы ядра. Женские половые гормоны ингибируют СМП и рост мышц у крыс (115), но не выявлено измеримых различий между мужчинами и женщинами в базальном уровне фракционного белкового синтеза или ответе на внутривенное введение аминокислот при умеренном уровне инсулина (Smith G., Mittendorfer B., Atherton P. и Rennie M. J., неопубликованные данные). Также не выявлены различия СМП или РМП в исходном уровне или после упражнений между молодыми мужчинами и женщинами (44, 59, 75, 82).
Тем не менее, согласно нашим и другим сообщениям, у женщин после менопаузы исходный уровень СМП на ~20 – 30 % выше, чем у мужчин (56, 105), и незначительно реагирует на прием пищи (105), таким образом, различия в метаболизме белка проявляются с возрастом, вероятно, в результате изменения в гормональном статусе. Изменения возникают независимо от композиции тела, так, нами были обследованы субъекты с ожирением (ИМТ 36–38) (105), в отличие от эксперимента группы Nair’s (56), сообщавшей об аналогичных различиях в базальном уровне СМП. Известно, что пожилые женщины демонстрируют меньшую гипертрофию (~33 %), чем мужчины, в ответ на тренировку с отягощением (3 дня в неделю, 26 недель) (6), вероятно, из-за неспособности поддерживать адаптивный ответ на хроническое тренирующее воздействие; так, пожилые мужчины увеличивают базальный уровень СМП на ~50 % после 3 месяцев тренировки, тогда как пожилые женщины только на ~15 % (104)
Влияние возраста на метаболизм мышечного белка и упражнения
 Противоречивы сведения относительно базального уровня СМП у пожилых людей, в некоторых ранних исследованиях сообщалось о снижении уровня синтеза белка в мышцах по сравнению с молодыми субъектами (96, 121, 127). Тем не менее, если данные корректны, величина утраты мышечной ткани с возрастом должна быть намного выше, чем обычно наблюдаемая, поэтому большинство ученых в настоящее время согласны, что у здоровых людей старение не влияет на базальный уровень СМП, и общий баланс белка у здоровых пожилых людей не снижается (32, 117, 119). Умеренное повышение двигательной активности предотвращает связанную с возрастом утрату мышечной силы, а также возрастную жировую инфильтрацию мышц у пожилых людей (40). Более того, было показано, что у пожилых людей СМП может стимулироваться как упражнениями с отягощением, так и питанием (32, 38, 131). Тем не менее мы недавно продемонстрировали «анаболическую резистентность» СМП стариков к однократной тренировке из упражнений с отягощениями в широком диапазоне интенсивности нагрузки – на ~30 % меньший ответ у пожилых в сравнении с молодыми людьми (66).
Противоречивы сведения относительно базального уровня СМП у пожилых людей, в некоторых ранних исследованиях сообщалось о снижении уровня синтеза белка в мышцах по сравнению с молодыми субъектами (96, 121, 127). Тем не менее, если данные корректны, величина утраты мышечной ткани с возрастом должна быть намного выше, чем обычно наблюдаемая, поэтому большинство ученых в настоящее время согласны, что у здоровых людей старение не влияет на базальный уровень СМП, и общий баланс белка у здоровых пожилых людей не снижается (32, 117, 119). Умеренное повышение двигательной активности предотвращает связанную с возрастом утрату мышечной силы, а также возрастную жировую инфильтрацию мышц у пожилых людей (40). Более того, было показано, что у пожилых людей СМП может стимулироваться как упражнениями с отягощением, так и питанием (32, 38, 131). Тем не менее мы недавно продемонстрировали «анаболическую резистентность» СМП стариков к однократной тренировке из упражнений с отягощениями в широком диапазоне интенсивности нагрузки – на ~30 % меньший ответ у пожилых в сравнении с молодыми людьми (66).
Тогда как другие исследователи сообщали о запаздывании ответной реакции на упражнения с отягощениями у пожилых людей (38), мы не обнаружили подобного эффекта (66). Эти расхождения могут быть вызваны меньшим объемом упражнений, использованных в эксперименте, и обстоятельством, что субъекты изучались в состоянии голода после ночного сна (66). Подобный «притупленный» анаболический эффект наблюдался у стариков в ответ на прием пищи, расцененный как снижение чувствительности и способности к анаболическому ответу на прием аминокислот отдельно (32) и в сочетании с глюкозой (118).
Худший анаболический ответ СМП на упражнения с отягощением в мышцах пожилых людей, видимо, связан со снижением активации восходящего сигнала пути mTOR и повышением активности АМРК по сравнению с мышцами молодых людей (38). В исследованиях на грызунах обнаружено снижение активации сигнального пути mTOR после упражнений с отягощениями и повышение активности АМРК у пожилых крыс по сравнению с молодыми животными (45, 83, 111).
Недостаточное количество данных об РМП в ответ на упражнения у стариков. Используя метод растворенных A-V меток, обнаружено некоторое увеличение распада белка в состоянии покоя у пожилых людей (119). Тем не менее, по нашим данным, нет различий в базальном РМП, но «нормальное» ингибирование РМП инсулином существенно ниже у стариков (122). Видимо, «притупленный анаболизм» – широко распространенное явление в стареющих мышцах.
Минимизация потерь мышечной ткани, поддержание массы и функций мышц – основная цель при старении; мы должны понять синергию между упражнениями и приемом пищи, а также разработать надлежащую стратегию тренировок и питания для пожилых людей.
Выводы
Подводя итог, можно констатировать, что мышцы показывают удивительную пластичность в ответ на упражнения. Во время упражнений с отягощениями и без них СМП подавляется, тогда как после тренировки – повышается как в голодном, так и в сытом состоянии. Эта стимуляция проявляет зависимость от дозы и величины воздействия; тем не менее, для выяснение роли рабочей нагрузки требуются дополнительные исследования. Независимо от различий в нагрузке и наличия (или отсутствия) отягощения, сократительная активность вызывает сходный срочный анаболический ответ в нетренированных мышцах. Однако после периода тренировок срочный ответ уменьшается и зависит от вида упражнений: с отягощением или без отягощения и стимулирует синтез белков миофибрилл (с отягощением) или митохондрий (без отягощения), вероятно, отражая адаптивные изменения в зависимости от вида упражнения. Активация РМП во время упражнения с отягощением или без остается невыясненной. Тем не менее существует достаточно данных, подтверждающих увеличение РМП после обоих видов упражнений. Общее увеличение мышечной массы (СМП – РМП) после упражнений проявляется только при повышении доступности аминокислот во время постнагрузочного периода. Приблизительно 20 г высококачественного белка, например молочного, достаточно для инициации максимального синтетического ответа и, соответственно, общего увеличения массы мышц.
С возрастом уменьшается синтез белка миофибрилл в ответ на упражнение и потребление пищи, последние данные свидетельствуют о существовании половых различий в обмене мышечного белка, особенно об уменьшении ответа на упражнения у пожилых женщин.
Активация АМРК в результате клеточных запасов энергии играет важную роль в ингибировании синтеза белка. Увеличение синтеза белка после упражнений опосредуется активацией передачи сигналов mTOR и последующих эффекторов. Связь времени и величины ответа между упражнениями и фосфорилированием регуляторов клеточных сигнальных путей, вовлеченных в синтез и деградацию белка, только начинает выясняться. Несмотря на то что эти процессы фосфорилирования в основном качественно проявляются в результате анаболических стимулов, таких как упражнения, требуется гораздо больше работ для расшифровки сигналов, которые их включают или выключают и в конечном счете контролируют адаптивный ответ, например, мышечную гипертрофию или биогенез митохондрий. Конечно, в настоящее время невозможно напрямую связать размеры активации обмена мышечных белков с фосфорилированием сигнальных молекул. Когда мы сможем сделать это, мы будем гораздо ближе к нашей цели – пониманию регуляции мышечной массы и функции и разработке стратегии максимизации поддержания мышц в здоровье и болезни.
Источник: https://journals.physiology.org/
Источники:
1.
Aronson D., Violan M.A., Dufresne S.D., Zangen D., Fielding R.A.,
Goodyear L.J. Exercise stimulates the mitogen-activated protein kinase pathway in human skeletal muscle. J ClinInvest, 99: 1251–1257, 1997.
Medline
2.
Atherton P.J., Babraj J., Smith K., Singh J., Rennie M.J., Wackerhage H. Selective activation of AMPK-PGC-1alpha or PKB-TSC2-mTOR signaling can explain specific adaptive responses to endurance or resistance training-like electrical muscle stimulation. FASEB J, 19: 786–788, 2005.
Abstract/FREE FullText
3.
Attaix D., Ventadour S., Codran A., Bechet D., Taillandier D., Combaret L.The ubiquitin-proteasome system and skeletal muscle wasting. Essays Biochem, 41: 173–186, 2005.
CrossRef Medline
4.
Baar K., Esser K. Phosphorylation of p70(S6k) correlates with increased skeletal muscle mass following resistance exercise. Am J Physiol Cell Physiol, 276: C120–C127, 1999.
Abstract/FREE Full Text
5.
Balagopal P., Schimke J.C., Ades P., Adey D., Nair K.S. Age effect on transcript levels and synthesis rate of muscle MHC and response to resistance exercise. Am J Physiol Endocrinol Metab, 280: E203–E208, 2001.
Abstract/FREE Full Text
6.
Bamman M.M., Hill V.J., Adams G.R., Haddad F., Wetzstein C.J., Gower B.A., Ahmed A., Hunter G.R. Gender differences in resistance-training-induced myofiber hypertrophy among older adults. J Gerontol A BiolSciMedSci, 58: 108–116, 2003.
Medline
7.
Beelen M., Koopman R., Gijsen A.P., Vandereyt H., Kies A.K., Kuipers H., Saris W.H., van Loon L.J. Protein coingestion stimulates muscle protein synthesis during resistance-type exercise. Am J Physiol Endocrinol Metab, 295: E70–E77, 2008.
Abstract/FREE Full Text
8.
Beelen M., Tieland M., Gijsen A.P., Vandereyt H., Kies A.K., Kuipers H., Saris W.H., Koopman R., van Loon L.J. Coingestion of carbohydrate and protein hydrolysate stimulates muscle protein synthesis during exercise in young men, with no further increase during subsequent overnight recovery. J Nutr, 138: 2198–2204, 2008.
Abstract/FREE FullText
9.
Belcastro A.N. Skeletal muscle calcium-activated neutral protease (calpain) with exercise .J ApplPhysiol, 74: 1381–1386, 1993.
Abstract/FREE FullText
10.
Bennet W.M., Connacher A.A., Scrimgeour C.M., Smith K., Rennie M.J. Increase in anterior tibialis muscle protein synthesis in healthy man during mixed amino acid infusion: studies of incorporation of [1‑13C]leucine. ClinSci (Lond), 76: 447–454, 1989.
Medline
11.
Benziane B., Burton T.J., Scanlan B., Galuska D., Canny B.J., Chibalin A.V., Zierath J.R., Stepto N.K. Divergent cell signaling after short-term intensified endurance training in human skeletal muscle. Am J Physiol Endocrinol Metab, 295: E1427–E1438, 2008.
Abstract/FREE Full Text
12.
Bier D.M.. Stable isotopes in biosciences, their measurement and models for amino acid metabolism. Eur J Pediatr, 156, Suppl 1: S2–S8, 1997.
CrossRef
Medline
13.
Biolo G., Maggi S.P., Williams B.D., Tipton K.D., Wolfe R.R. Increased rates of muscle protein turnover and amino acid transport after resistance exercise in humans. Am J Physiol Endocrinol Metab, 268: E514–E520, 1995.
Abstract/FREE Full Text
14.
Biolo G., Tipton K.D., Klein S., Wolfe R.R. An abundant supply of amino acids enhances the metabolic effect of exercise on muscle protein. Am J Physiol Endocrinol Metab, 273: E122–E129, 1997.
Abstract/FREE Full Text
15.
Biolo G., Williams B.D., Fleming R.Y., Wolfe R.R. Insulin action on muscle protein kinetics and amino acid transport during recovery after resistance exercise. Diabetes, 48: 949–957, 1999.
Abstract
16.
Bohe J., Low A., Wolfe R.R., Rennie M.J. Human muscle protein synthesis is modulated by extracellular, not intramuscular amino acid availability: a dose-response study. J Physiol, 552: 315–324, 2003.
Abstract/FREE FullText
17.
Bohe J, Low J.F., Wolfe R.R., Rennie M.J. Latency and duration of stimulation of human muscle protein synthesis during continuous infusion of amino acids. J Physiol, 532: 575–579, 2001.
Abstract/FREE FullText
18.
Bolster D.R., Crozier S.J., Kimball S.R., Jefferson L.S. AMP-activated protein kinase suppresses protein synthesis in rat skeletal muscle through down-regulated mammalian target of rapamycin (mTOR) signaling. J BiolChem, 277: 23977–23980, 2002.
Abstract/FREE FullText
19.
Bolster D.R., Kubica N., Crozier S.J., Williamson D.L., Farrell P.A., Kimball S.R., Jefferson L.S. Immediate response of mammalian target of rapamycin (mTOR)-mediated signaling following acute resistance exercise in rat skeletal muscle. J Physiol, 553: 213–220, 2003.
Abstract/FREE FullText
20.
Borsheim E., Cree M.G., Tipton K.D., Elliott T.A., Aarsland A., Wolfe R.R. Effect of carbohydrate intake on net muscle protein synthesis during recovery from resistance exercise. J ApplPhysiol, 96: 674–678, 2004.
Abstract/FREE FullText
21.
Borsheim E., Tipton K.D., Wolf S.E., Wolfe R.R. Essential amino acids and muscle protein recovery from resistance exercise. Am J Physiol Endocrinol Metab, 283: E648–E657, 2002.
Abstract/FREE Full Text
22.
Bowtell J.L., Leese G.P., Smith K., Watt P.W., Nevill A., Rooyackers O., Wagenmakers A.J., Rennie M.J. Modulation of whole body protein metabolism, during and after exercise, by variation of dietary protein. J ApplPhysiol, 85: 1744–1752, 1998.
Abstract/FREE FullText
23.
Brodsky I.G., Balagopal P., Nair K.S. Effects of testosterone replacement on muscle mass and muscle protein synthesis in hypogonadal men – a clinical research center study. J Clin Endocrinol Metab, 81: 3469–3475, 1996.
Abstract
24.
Bylund-Fellenius A.C., Ojamaa K.M., Flaim K.E., Li J.B., Wassner S.J., Jefferson L.S. Protein synthesis versus energy state in contracting muscles of perfused rat hindlimb. Am J Physiol Endocrinol Metab, 246: E297–E305, 1984.
Abstract/FREE Full Text
25.
Carmeli E., Moas M., Lennon S., Powers S.K. High intensity exercise increases expression of matrix metalloproteinases in fast skeletal muscle fibres. ExpPhysiol, 90: 613–619, 2005.
Abstract/FREE FullText
26.
Carraro F., Hartl W.H., Stuart C.A., Layman D.K., Jahoor F., Wolfe R.R. Whole body and plasma protein synthesis in exercise and recovery in human subjects. Am J Physiol Endocrinol Metab, 258: E821–E831, 1990.
Abstract/FREE Full Text
27.
Carraro F., Stuart C.A., Hartl W.H., Rosenblatt J., Wolfe R.R. Effect of exercise and recovery on muscle protein synthesis in human subjects. Am J Physiol Endocrinol Metab, 259: E470–E476, 1990.
Abstract/FREE Full Text
28.
Cheng K.N., Dworzak F., Ford G.C., Rennie M.J., Halliday D. Direct determination of leucine metabolism and protein breakdown in humans using l-[1-13C,15N]-leucine and the forearm model. Eur J ClinInvest, 15: 349–354, 1985.
Medline
29.
Chesley A., MacDougall J.D., Tarnopolsky M.A., Atkinson S.A., Smith K. Changes in human muscle protein synthesis after resistance exercise. J ApplPhysiol, 73: 1383–1388, 1992.
Abstract/FREE FullText
30.
Chinkes D., Klein S., Zhang X.J., Wolfe R.R. Infusion of labeled KIC is more accurate than labeled leucine to determine human muscle protein synthesis. Am J Physiol Endocrinol Metab, 270: E67–E71, 1996.
Abstract/FREE Full Text
31.
Coffey V.G., Hawley J.A. The molecular bases of training adaptation. SportsMed, 37: 737–763, 2007.
CrossRef Medline
32.
Cuthbertson D., Smith K., Babraj J., Leese G., Waddell T., Atherton P., Wackerhage H., Taylor P.M., Rennie M.J. Anabolic signaling deficits underlie amino acid resistance of wasting, aging muscle. FASEB J, 19: 422–424, 2005.
Abstract/FREE FullText
33.
Cuthbertson D.J., Babraj J., Smith K., Wilkes E., Fedele M.J., Esser K., Rennie M. Anabolic signaling and protein synthesis in human skeletal muscle after dynamic shortening or lengthening exercise. Am J Physiol Endocrinol Metab,290: E731–E738, 2006.
Abstract/FREE Full Text
34.
Deldicque L., Atherton P., Patel R., Theisen D., Nielens H., Rennie M.J., Francaux M. Decrease in Akt/PKB signalling in human skeletal muscle by resistance exercise. Eur J ApplPhysiol, 104: 57–65, 2008.
CrossRef Medline
35.
Dohm G.L., Tapscott E.B., Barakat H.A., Kasperek G.J. Measurement of in vivo protein synthesis in rats during an exercise bout. BiochemMed, 27: 367–373, 1982.
CrossRef Medline
36.
Dreyer H.C., Drummond M.J., Pennings B., Fujita S., Glynn E.L., Chinkes D.L., Dhanani S., Volpi E., Rasmussen B.B. Leucine-enriched essential amino acid and carbohydrate ingestion following resistance exercise enhances mTOR signaling and protein synthesis in human muscle. Am J Physiol Endocrinol Metab, 294: E392–E400, 2008.
Abstract/FREE Full Text
37.
Dreyer H.C., Fujita S., Cadenas J.G., Chinkes D.L., Volpi E., Rasmussen B.B. Resistance exercise increases AMPK activity and reduces 4E-BP1 phosphorylation and protein synthesis in human skeletal muscle. J Physiol, 576: 613–624, 2006.
Abstract/FREE FullText
38.
Drummond M.J., Dreyer H.C., Pennings B., Fry C.S., Dhanani S., Dillon E.L., Sheffield-Moore M., Volpi E., Rasmussen B.B. Skeletal muscle protein anabolic response to resistance exercise and essential amino acids is delayed with aging. J ApplPhysiol, 104: 1452–1461, 2008.
Abstract/FREE FullText
39.
Durham W.J., Miller S.L., Yeckel C.W., Chinkes D.L., Tipton K.D., Rasmussen B.B., Wolfe R.R. Leg glucose and protein metabolism during an acute bout of resistance exercise in humans. J ApplPhysiol, 97: 1379–1386, 2004.
Abstract/FREE FullText
40.
Esmarck B., Andersen J.L., Olsen S., Richter E.A., Mizuno M., Kjaer M. Timing of postexercise protein intake is important for muscle hypertrophy with resistance training in elderly humans. J Physiol, 535: 301–311, 2001.
Abstract/FREE FullText
41.
Forbes G.B. The effect of anabolic steroids on lean body mass: the dose response curve. Metabolism, 34: 571–573, 1985.
CrossRef Medline
42.
Fujii N., Hayashi T., Hirshman M.F., Smith J.T., Habinowski S.A., Kaijser L., Mu J., Ljungqvist O., Birnbaum M.J., Witters L.A., Thorell A., Goodyear L.J. Exercise induces isoform-specific increase in 5′AMP-activated protein kinase activity in human skeletal muscle. BiochemBiophysResCommun, 273: 1150–1155, 2000.
CrossRef
Medline
43.
Fujita S., Dreyer H.C., Drummond M.J., Glynn E.L., Volpi E., Rasmussen B.B. Essential amino acid and carbohydrate ingestion prior to resistance exercise does not enhance post-exercise muscle protein synthesis. J ApplPhysiol. Inpress.
44.
Fujita S., Rasmussen B.B., Bell J.A., Cadenas J.G., Volpi E. Basal muscle intracellular amino acid kinetics in women and men. Am J Physiol Endocrinol Metab, 292: E77–E83, 2007.
Abstract/FREE Full Text
45.
Funai K., Parkington J.D., Carambula S., Fielding R.A. Age-associated decrease in contraction-induced activation of downstream targets of Akt/mTor signaling in skeletal muscle. Am J Physiol Regul Integr Comp Physiol, 290: R1080–R1086, 2006.
Abstract/FREE Full Text
46.
Gautsch T.A., Anthony J.C., Kimball S.R., Paul G.L., Layman D.K., Jefferson L.S. Availability of eIF4E regulates skeletal muscle protein synthesis during recovery from exercise. Am J Physiol Cell Physiol, 274: C406–C414, 1998.
Abstract/FREE Full Text
47.
Glover E.I., Phillips S.M., Oates B.R., Tang J.E., Tarnopolsky M.A., Selby A., Smith K., Rennie M.J. Immobilization induces anabolic resistance in human myofibrillar protein synthesis with low and high dose amino acid infusion. J Physiol, 586: 6049–61, 2008.
Abstract/FREE FullText
48.
Gollnick P.D., Saltin B. Significance of skeletal muscle oxidative enzyme enhancement with endurance training. ClinPhysiol, 2: 1–12, 1982.
Medline
49.
Goodpaster B.H., Chomentowski P., Ward B.K., Rossi A., Glynn N.W., Delmonico M.J., Kritchevsky S.B., Pahor M., Newman A.B. Effects of physical activity on strength and skeletal muscle fat infiltration in older adults: a randomized controlled trial. J ApplPhysiol, 105: 1498–1503, 2008.
Abstract/FREE FullText
50.
Greenhaff P.L., Karagounis L., Peirce N., Simpson E.J., Hazell M., Layfield R., Wackerhage H., Smith K., Atherton P., Selby A., Rennie M.J. Disassociation between the effects of amino acids and insulin on signaling, ubiquitin-ligases and protein turnover in human muscle. Am J Physiol Endocrinol Metab, 295: E595–E604, 2008.
Abstract/FREE Full Text
51.
Halliday D., McKeran R.O. Measurement of muscle protein synthetic rate from serial muscle biopsies and total body protein turnover in man by continuous intravenous infusion of l-(α-15N)lysine. ClinSciMolMed, 49: 581–590, 1975.
Medline
52.
Hardie D.G. AMPK: a key regulator of energy balance in the single cell and the whole organism. Int J Obes (Lond), 32, Suppl4: S7–S12, 2008.
53.
Hartman J.W., Tang J.E., Wilkinson S.B., Tarnopolsky M.A., Lawrence R.L., Fullerton A.V., Phillips S.M. Consumption of fat-free fluid milk after resistance exercise promotes greater lean mass accretion than does consumption of soy or carbohydrate in young, novice, male weightlifters. Am J ClinNutr, 86: 373–381, 2007.
Abstract/FREE FullText
54.
Hasten D.L., Pak-Loduca J., Obert K.A., Yarasheski K.E. Resistance exercise acutely increases MHC and mixed muscle protein synthesis rates in 78–84 and 23–32 yr olds. Am J Physiol Endocrinol Metab, 278: E620–E626, 2000.
Abstract/FREE Full Text
55.
Haus J.M., Miller B.F., Carroll C.C., Weinheimer E.M., Trappe T.A. The effect of strenuous aerobic exercise on skeletal muscle myofibrillar proteolysis in humans. Scand J MedSciSports, 17: 260–266, 2007.
Medline
56.
Henderson G.C., Dhatariya K., Ford G.C., Klaus K.A., Basu R., Rizza R.A., Jensen M.D., Khosla S., O’Brien P., Nair K.S. Higher muscle protein synthesis in women than men across the lifespan, and failure of androgen administration to amend age-related decrements. FASEB J, 23: 631–641, 2009.
Abstract/FREE FullText
57.
Hoppeler H. Exercise-induced ultrastructural changes in skeletal muscle. Int J SportsMed, 7: 187–204, 1986.
Medline
58.
Isidori A.M., Giannetta E., Greco E.A., Gianfrilli D., Bonifacio V., Isidori A., Lenzi A., Fabbri A. Effects of testosterone on body composition, bone metabolism and serum lipid profile in middle-aged men: a meta-analysis. ClinEndocrinol (Oxf), 63: 280–293, 2005.
CrossRef Medline
59.
Jahn L.A., Barrett E.J., Genco M.L., Wei L., Spraggins T.A., Fryburg D.A. Tissue composition affects measures of postabsorptive human skeletal muscle metabolism: comparison across genders. J ClinEndocrinolMetab, 84: 1007–1010, 1999.
Abstract/FREE FullText
60.
Jaleel A., Short K.R., Asmann Y.W., Klaus K.A., Morse D.M., Ford G.C., Nair K.S. In vivo measurement of synthesis rate of individual skeletal muscle mitochondrial proteins. Am J Physiol Endocrinol Metab, 295: E1255–E1268, 2008.
Abstract/FREE Full Text
61.
Karlsson H.K., Nilsson P.A., Nilsson J., Chibalin A.V., Zierath J.R., Blomstrand E. Branched-chain amino acids increase p70S6k phosphorylation in human skeletal muscle after resistance exercise. Am J Physiol Endocrinol Metab, 287: E1–E7, 2004.
Abstract/FREE Full Text
62.
Kim P.L., Staron R.S., Phillips S.M. Fasted-state skeletal muscle protein synthesis after resistance exercise is altered with training. J Physiol, 568: 283–290, 2005.
Abstract/FREE FullText
63.
Kimball S.R., Farrell P.A., Jefferson L.S. Invited Review: Role of insulin in translational control of protein synthesis in skeletal muscle by amino acids or exercise. J ApplPhysiol, 93: 1168–1180, 2002.
Abstract/FREE FullText
64.
Koopman R., Wagenmakers A.J., Manders R.J., Zorenc A.H., Senden J.M., Gorselink M., Keizer H.A., van Loon L.J. Combined ingestion of protein and free leucine with carbohydrate increases postexercise muscle protein synthesis in vivo in male subjects. Am J Physiol Endocrinol Metab, 288: E645–E653, 2005.
Abstract/FREE Full Text
65.
Kostek M.C., Chen Y.W., Cuthbertson D.J., Shi R., Fedele M.J., Esser K.A., Rennie M.J. Gene expression responses over 24 h to lengthening and shortening contractions in human muscle: major changes in CSRP3, MUSTN1, SIX1, and FBXO32. PhysiolGenomics, 31: 42–52, 2007.
Abstract/FREE FullText
66.
Kumar V., Selby A., Rankin D., Patel R., Atherton P., Hildebrandt W., Williams J., Smith K., Seynnes O., Hiscock N., Rennie M.J. Age-related differences in the dose-response of muscle protein synthesis to resistance exercise in young and old men. J Physiol, 587: 211–217, 2009.
Abstract/FREE FullText
67.
Levenhagen D.K., Gresham J.D., Carlson M.G., Maron D.J., Borel M.J., Flakoll P.J. Postexercise nutrient intake timing in humans is critical to recovery of leg glucose and protein homeostasis. Am J Physiol Endocrinol Metab, 280: E982–E993, 2001.
Abstract/FREE Full Text
68.
Louard R.J., Barrett E.J., Gelfand R.A. Effect of infused branched-chain amino acids on muscle and whole-body amino acid metabolism in man. ClinSci (Lond), 79: 457–466, 1990.
Medline
69.
Louard R.J., Barrett E.J., Gelfand R.A. Overnight branched-chain amino acid infusion causes sustained suppression of muscle proteolysis. Metabolism, 44: 424–429, 1995.
CrossRef Medline
70.
Louis M., Poortmans J.R., Francaux M., Berre J., Boisseau N., Brassine E., Cuthbertson D.J., Smith K., Babraj J.A., Waddell T., Rennie M.J. No effect of creatine supplementation on human myofibrillar and sarcoplasmic protein synthesis after resistance exercise. Am J Physiol Endocrinol Metab, 285: E1089–E1094, 2003.
Abstract/FREE Full Text
71.
MacDougall J.D., Gibala M.J., Tarnopolsky M.A., MacDonald J.R., Interisano S.A., Yarasheski K.E. The time course for elevated muscle protein synthesis following heavy resistance exercise. Can J ApplPhysiol, 20: 480–486, 1995.
Medline
72.
MacLean D.A., Graham T.E., Saltin B. Branched-chain amino acids augment ammonia metabolism while attenuating protein breakdown during exercise. Am J Physiol Endocrinol Metab, 267: E1010–E1022, 1994.
Abstract/FREE Full Text
73.
MacLean D.A., Graham T.E., Saltin B. Stimulation of muscle ammonia production during exercise following branched-chain amino acid supplementation in humans. J Physiol, 493: 909–922, 1996.
Abstract/FREE FullText
74.
Mascher H., Andersson H., Nilsson P.A., Ekblom B., Blomstrand E. Changes in signalling pathways regulating protein synthesis in human muscle in the recovery period after endurance exercise. ActaPhysiol (Oxf), 191: 67–75, 2007.
CrossRef Medline
75.
Miller B.F., Hansen M., Olesen J.L., Flyvbjerg A., Schwarz P., Babraj J.A., Smith K., Rennie M.J., Kjaer M. No effect of menstrual cycle on myofibrillar and connective tissue protein synthesis in contracting skeletal muscle. Am J Physiol Endocrinol Metab, 290: E163–E168, 2006.
Abstract/FREE Full Text
76.
Miller B.F., Olesen J.L., Hansen M., Dossing S., Crameri R.M., Welling R.J., Langberg H., Flyvbjerg A., Kjaer M., Babraj J.A., Smith K., Rennie M.J. Coordinated collagen and muscle protein synthesis in human patella tendon and quadriceps muscle after exercise. J Physiol, 567: 1021–1033, 2005.
Abstract/FREE FullText
77.
Miller S.L., Tipton K.D., Chinkes D.L., Wolf S.E., Wolfe R.R. Independent and combined effects of amino acids and glucose after resistance exercise. MedSciSportsExerc, 35: 449–455, 2003.
CrossRef Medline
78.
Millward D.J., Jackson A.A. Protein/energy ratios of current diets in developed and developing countries compared with a safe protein/energy ratio: implications for recommended protein and amino acid intakes. PublicHealthNutr, 7: 387–405, 2004.
CrossRef Medline
79.
Moore D.R., Phillips S.M., Babraj J.A., Smith K., Rennie M.J. Myofibrillar and collagen protein synthesis in human skeletal muscle in young men after maximal shortening and lengthening contractions. Am J Physiol Endocrinol Metab, 288: E1153–E1159, 2005.
Abstract/FREE Full Text
80.
Moore D.R., Robinson M.J., Fry J.L., Tang J.E., Glover E.I., Wilkinson S.B., Prior T., Tarnopolsky M.A., Phillips S.M. Ingested protein dose-response of muscle and albumin prtoein synthesis after resistance exercise in young men. Am J ClinNutr, 89: 161–168, 2008.
CrossRef Medline
81.
Nair K.S., Jaleel A., Asmann Y.W., Short K.R., Raghavakaimal S. Proteomic research: potential opportunities for clinical and physiological investigators. Am J Physiol Endocrinol Metab, 286: E863–E874, 2004.
Abstract/FREE Full Text
82.
Parise G., Mihic S., MacLennan D., Yarasheski K.E., Tarnopolsky M.A. Effects of acute creatine monohydrate supplementation on leucine kinetics and mixed-muscle protein synthesis. J ApplPhysiol, 91: 1041–1047, 2001.
Abstract/FREE FullText
83.
Parkington J.D., LeBrasseur N.K., Siebert A.P., Fielding R.A. Contraction-mediated mTOR, p70S6k, and ERK1/2 phosphorylation in aged skeletal muscle. J ApplPhysiol, 97: 243–248, 2004.
Abstract/FREE FullText
84.
Phillips S.M., Parise G., Roy B.D., Tipton K.D., Wolfe R.R., Tamopolsky M.A. Resistance-training-induced adaptations in skeletal muscle protein turnover in the fed state. Can J PhysiolPharmacol, 80: 1045–1053, 2002.
CrossRef
Medline
85.
Phillips S.M., Tipton K.D., Aarsland A., Wolf S.E., Wolfe R.R. Mixed muscle protein synthesis and breakdown after resistance exercise in humans. Am J Physiol Endocrinol Metab, 273: E99–E107, 1997.
Abstract/FREE Full Text
86.
Phillips S.M., Tipton K.D., Ferrando A.A., Wolfe R.R. Resistance training reduces the acute exercise-induced increase in muscle protein turnover. Am J Physiol Endocrinol Metab, 276: E118–E124, 1999.
Abstract/FREE Full Text
87.
Pikosky M.A., Gaine P.C., Martin W.F., Grabarz K.C., Ferrando A.A., Wolfe R.R., Rodriguez N.R. Aerobic exercise training increases skeletal muscle protein turnover in healthy adults at rest. J Nutr, 136: 379–383, 2006.
Abstract/FREE FullText
88.
Pilegaard H., Richter E.A. PGC-1alpha: important for exercise performance? J ApplPhysiol, 104: 1264–1265, 2008.
FREE FullText
89.
Pilegaard H., Saltin B., Neufer P.D. Exercise induces transient transcriptional activation of the PGC-1alpha gene in human skeletal muscle. J Physiol, 546: 851–858, 2003.
Abstract/FREE FullText
90.
Rasmussen B.B., Tipton K.D., Miller S.L., Wolf S.E., Wolfe R.R. An oral essential amino acid-carbohydrate supplement enhances muscle protein anabolism after resistance exercise. J ApplPhysiol, 88: 386–392, 2000.
Abstract/FREE FullText
91.
Raue U., Slivka D., Jemiolo B., Hollon C., Trappe S. Proteolytic gene expression differs at rest and after resistance exercise between young and old women. J Gerontol A Biol Sci Med Sci, 62: 1407–1412, 2007.
Abstract/FREE Full Text
92.
Rennie M.J. An introduction to the use of tracers in nutrition and metabolism.ProcNutrSoc, 58: 935–944, 1999.
Medline
93.
Rennie M.J., Edwards R.H., Davies C.T., Krywawych S., Halliday D., Waterlow J.C., Millward D.J. Protein and amino acid turnover during and after exercise. Biochem Soc Trans, 8: 499–501, 1980.
Medline
94.
Rennie M.J., Edwards R.H., Halliday D., Matthews D.E., Wolman S.L., Millward D.J. Muscle protein synthesis measured by stable isotope techniques in man: the effects of feeding and fasting. ClinSci (Lond), 63: 519–523, 1982.
Medline
95.
Rennie M.J., Phillips S., Smith K. Reliability of results and interpretation of measures of 3-methylhistidine in muscle interstitium as marker of muscle proteolysis. J ApplPhysiol, 105: 1380–1381, 2008.
FREE FullText
96.
Rooyackers O.E., Adey D.B., Ades P.A., Nair K.S. Effect of age on in vivo rates of mitochondrial protein synthesis in human skeletal muscle. ProcNatlAcadSci USA, 93: 15364–15369, 1996.
Abstract/FREE FullText
97.
Rose A.J., Broholm C., Kiillerich K., Finn S.G., Proud C.G., Rider M.H., Richter E.A., Kiens B. Exercise rapidly increases eukaryotic elongation factor 2 phosphorylation in skeletal muscle of men. J Physiol, 569: 223–228, 2005.
Abstract/FREE FullText
98.
Sakamoto K., Arnolds D.E., Ekberg I., Thorell A., Goodyear L.J. Exercise regulates Akt and glycogen synthase kinase-3 activities in human skeletal muscle. BiochemBiophysResCommun, 319: 419–425, 2004.
CrossRef Medline
99.
Sakamoto K., Goodyear L.J. Invited Review: Intracellular signaling in contracting skeletal muscle. J ApplPhysiol, 93: 369–383, 2002.
Abstract/FREE FullText
100.
Sheffield-Moore M., Paddon-Jones D., Sanford A.P., Rosenblatt J.I., Matlock A.G., Cree M.G., Wolfe R.R. Mixed muscle and hepatic derived plasma protein metabolism is differentially regulated in older and younger men following resistance exercise. Am J Physiol Endocrinol Metab, 288: E922–E929, 2005.
Abstract/FREE Full Text
101.
Sheffield-Moore M., Yeckel C.W., Volpi E., Wolf S.E., Morio B., Chinkes D.L., Paddon-Jones D., Wolfe R.R. Postexercise protein metabolism in older and younger men following moderate-intensity aerobic exercise. Am J Physiol Endocrinol Metab, 287: E513–E522, 2004.
Abstract/FREE Full Text
102.
Short K.R., Vittone J.L., Bigelow M.L., Proctor D.N., Nair K.S. Age and aerobic exercise training effects on whole body and muscle protein metabolism. Am J Physiol Endocrinol Metab, 286: E92–E101, 2004.
Abstract/FREE Full Text
103.
Short K.R., Vittone J.L., Bigelow M.L., Proctor D.N., Rizza R.A., Coenen-Schimke J.M., Nair K.S. Impact of aerobic exercise training on age-related changes in insulin sensitivity and muscle oxidative capacity. Diabetes, 52: 1888–1896, 2003.
Abstract/FREE FullText
104.
Smith G., Villareal D., Sinacore D., Shah K., Mittendorfer B. The anabolic response to exercise training is greater in older men than older women. In: APS Intersociety Meeting: The Integrative Biology of Exercise V, September 24–27, 2009, Hilton Head, SC. Bethesda, MD: Am. Physiol. Soc., 2008, p. 17.2.
105.
Smith G.I., Atherton P., Villareal D.T., Frimel T.N., Rankin D., Rennie M.J., Mittendorfer B. Differences in muscle protein synthesis and anabolic signaling in the postabsorptive state and in response to food in 65–80 year old men and women. PLoS ONE3: e1875, 2008.
CrossRef
Medline
106.
Smith K., Barua J.M., Watt P.W., Scrimgeour C.M., Rennie M.J. Flooding with l-[1-13C]leucine stimulates human muscle protein incorporation of continuously infused l-[1-13C]valine. Am J Physiol Endocrinol Metab, 262: E372–E376, 1992.
Abstract/FREE Full Text
107.
Smith K., Rennie M.J. The measurement of tissue protein turnover. BaillieresClinEndocrinolMetab, 10: 469–495, 1996.
CrossRef
Medline
108.
Tang J.E., Manolakos J.J., Kujbida G.W., Lysecki P.J., Moore D.R., Phillips S.M. Minimal whey protein with carbohydrate stimulates muscle protein synthesis following resistance exercise in trained young men. ApplPhysiolNutrMetab, 32: 1132–1138, 2007.
CrossRef
Medline
109.
Tang J.E., Perco J.G., Moore D.R., Wilkinson S.B., Phillips S.M. Resistance training alters the response of fed state mixed muscle protein synthesis in young men. Am J Physiol Regul Integr Comp Physiol, 294: R172–R178, 2008.
Abstract/FREE Full Text
110.
Thomson D.M., Gordon S.E. Diminished overload-induced hypertrophy in aged fast-twitch skeletal muscle is associated with AMPK hyperphosphorylation. J ApplPhysiol, 98: 557–564, 2005.
Abstract/FREE FullText
111.
Tipton K.D., Borsheim E., Wolf S.E., Sanford A.P., Wolfe R.R. Acute response of net muscle protein balance reflects 24-h balance after exercise and amino acid ingestion. Am J Physiol Endocrinol Metab, 284: E76–E89, 2003.
Abstract/FREE Full Text
112.
Tipton K.D., Ferrando A.A., Phillips S.M., Doyle D. Jr., Wolfe R.R. Postexercise net protein synthesis in human muscle from orally administered amino acids. Am J Physiol Endocrinol Metab, 276: E628–E634, 1999.
Abstract/FREE Full Text
113.
Tipton K.D., Ferrando A.A., Williams B.D., Wolfe R.R. Muscle protein metabolism in female swimmers after a combination of resistance and endurance exercise. J ApplPhysiol, 81: 2034–2038, 1996.
Abstract/FREE FullText
114.
Tipton K.D., Rasmussen B.B., Miller S.L., Wolf S.E., Owens-Stovall S.K., Petrini B.E., Wolfe R.R. Timing of amino acid-carbohydrate ingestion alters anabolic response of muscle to resistance exercise. Am J Physiol Endocrinol Metab, 281: E197–E206, 2001.
Abstract/FREE Full Text
115.
Toth M.J., Poehlman E.T., Matthews D.E., Tchernof A., MacCoss M.J. Effects of estradiol and progesterone on body composition, protein synthesis, and lipoprotein lipase in rats. Am J Physiol Endocrinol Metab, 280: E496–E501, 2001.
Abstract/FREE Full Text
116.
Urban R.J., Bodenburg Y.H., Gilkison C., Foxworth J., Coggan A.R., Wolfe R.R., Ferrando A. Testosterone administration to elderly men increases skeletal muscle strength and protein synthesis. Am J Physiol Endocrinol Metab, 269: E820–E826, 1995.
Abstract/FREE Full Text
117.
Volpi E., Ferrando A.A., Yeckel C.W., Tipton K.D., Wolfe R.R. Exogenous amino acids stimulate net muscle protein synthesis in the elderly. J ClinInvest, 101: 2000–2007, 1998.
Medline
118.
Volpi E., Mittendorfer B., Rasmussen B.B., Wolfe R.R. The response of muscle protein anabolism to combined hyperaminoacidemia and glucose-induced hyperinsulinemia is impaired in the elderly. J ClinEndocrinolMetab, 85: 4481–4490, 2000.
Abstract/FREE FullText
119.
Volpi E., Sheffield-Moore M., Rasmussen B.B., Wolfe R.R. Basal muscle amino acid kinetics and protein synthesis in healthy young and older men. JAMA, 286: 1206–1212, 2001.
Abstract/FREE FullText
120.
Wagenmakers A.J. Tracers to investigate protein and amino acid metabolism in human subjects. ProcNutrSoc, 58: 987–1000, 1999.
Medline
121.
Welle S., Thornton C., Jozefowicz R., Statt M. Myofibrillar protein synthesis in young and old men. Am J Physiol Endocrinol Metab, 264: E693–E698, 1993.
Abstract/FREE Full Text
122.
Wilkes E., Selby A., Patel R., Rankin D., Smith K., Rennie M. Blunting of insulin-mediated proteolysis in leg muscle of elderly subjects may contribute to age-related sarcopenia (Abstract). ProcNutrSociety, 67 (OCE5): E153, 2008.
CrossRef
123.
Wilkinson S.B., Phillips S.M., Atherton P.J., Patel R., Yarasheski K.E., Tarnopolsky M.A., Rennie M.J. Differential effects of resistance and endurance exercise in the fed state on signaling molecule phosphorylation and protein synthesis in human muscle. J Physiol, 586: 3701–3717, 2008.
Abstract/FREE FullText
124.
Wilkinson S.B., Tarnopolsky M.A., Macdonald M.J., MacDonald J.R., Armstrong D., Phillips S.M. Consumption of fluid skim milk promotes greater muscle protein accretion after resistance exercise than does consumption of an isonitrogenous and isoenergetic soy-protein beverage. Am J ClinNutr, 85: 1031–1040, 2007.
Abstract/FREE FullText
125.
Williamson D.L., Kubica N., Kimball S.R., Jefferson L.S. Exercise-induced alterations in extracellular signal-regulated kinase 1/2 and mammalian target of rapamycin (mTOR) signalling to regulatory mechanisms of mRNA translation in mouse muscle. J Physiol, 573: 497–510, 2006.
Abstract/FREE FullText
126.
Yang Y., Creer A., Jemiolo B., Trappe S. Time course of myogenic and metabolic gene expression in response to acute exercise in human skeletal muscle. J ApplPhysiol, 98: 1745–1752, 2005.
Abstract/FREE FullText
127.
Yarasheski K.E. Exercise, aging, and muscle protein metabolism. J Gerontol A Biol Sci Med Sci, 58: M918–M922, 2003.
Abstract/FREE FullText
128.
Yarasheski K.E., Campbell J.A., Smith K., Rennie M.J., Holloszy J.O., Bier D.M. Effect of growth hormone and resistance exercise on muscle growth in young men. Am J Physiol Endocrinol Metab, 262: E261–E267, 1992.
Abstract/FREE Full Text
129.
Yarasheski K.E., Pak-Loduca J., Hasten D.L., Obert K.A., Brown M.B., Sinacore D.R. Resistance exercise training increases mixed muscle protein synthesis rate in frail women and men ≥76 yr old. Am J Physiol Endocrinol Metab, 277: E118–E125, 1999.
Abstract/FREE Full Text
130.
Yarasheski K.E., Smith K., Rennie M.J., Bier D.M. Measurement of muscle protein fractional synthetic rate by capillary gas chromatography/combustion isotope ratio mass spectrometry. BiolMassSpectrom, 21: 486–490, 1992.
CrossRef
Medline
131.
Yarasheski K.E., Zachwieja J.J., Bier D.M. Acute effects of resistance exercise on muscle protein synthesis rate in young and elderly men and women. Am J Physiol Endocrinol Metab, 265: E210–E214, 1993.
Abstract/FREE Full Text
132.
Zhang X.J., Chinkes D.L., Wolfe R.R. Measurement of muscle protein fractional synthesis and breakdown rates from a pulse tracer injection. Am J Physiol Endocrinol Metab, 283: E753–E764, 2002.
Abstract/FREE Full Text
Показать еще
Утомление и восстановление при мышечной деятельности
Физические
нагрузки и восстановление необходимо
рассматривать как две стороны процесса
повышения физической работоспособности
человека. Увеличение скорости
восстановительных процессов является
одним из основных показателей роста
тренированности спортсмена. Скорость
восстановительных процессов повышается
как за счет оптимизации режима нагрузок
и отдыха, так и с помощью вспомогательных
восстановительных средств.
При
этом различают два основные направления
воздействия на восстановительные
процессы:
1) повышение скорости
восстановления спортивной работоспособности
после тренировочных и соревновательных
нагрузок;
2) ускорение восстановительных
процессов после перенесенных заболеваний
и травм (реабилитационные меры).
Рациональное
использование различных восстановительных
средств позволяет повысить эффективность
тренировочного процесса. Однако в
некоторые периоды подготовки целесообразно
тренироваться в состоянии неполного
восстановления. Это стимулирует
адаптационные перестройки в организме,
позволяет в итоге подняться на более
высокий уровень работоспособности.
Восстановительные
средства принято делить на педагогические,
психологические и медико-биологические.
Главными считаются педагогические
средства восстановления. К ним относятся
рациональное чередование нагрузок и
отдыха, оптимальное соотношение объема
и интенсивности нагрузок, рациональное
планирование восстановительных циклов
подготовки и др.
Рациональному
использованию педагогических средств
будет способствовать рассмотрение
восстановительных процессов после
мышечной деятельности.
Характеристика
восстановительных процессов. При
выполнении физических упражнений в
организме расщепляются и восстанавливаются
богатые энергией соединения, причем
более интенсивно идут процессы
расщепления. Восстановительные же
процессы происходят в основном по
окончании работы. После рабочее состояние
организма называется восстановительным
периодом. Различают срочное (ранний
период) и отставленное восстановление
(поздний период).
Срочное
восстановление— это процесс
устранения накопившихся во время
выполнения упражнений продуктов обмена
в тканях (метаболитов) и ликвидация
образовавшегося кислородного долга.
Оно происходит непосредственно после
окончания каждого упражнения и
продолжается в течение 3090 минут после
выполнения тренировочной работы.
Отставленное
восстановление— возвращение к
исходному уровню энергетических ресурсов
организма, усиление синтеза структурных
белков и ферментов. Оно распространяется
на многие часы отдыха после работы.
Восстановление
разных функций организма происходит
неодновременно (гетерохронно). Показатели
скорости восстановления функций или
двигательных способностей после той
или иной работы используются при
планировании тренировочных нагрузок
с целью повышения эффекта от них. Быстрее
всего восстанавливаются резервы
фосфогенов и кислорода в работающих
мышцах, после них — запасы гликогена в
мышцах, затем — жировые резервы и
разрушенные белковые структуры клеток.
Интенсивность
протекания восстановительных процессов
определяется скоростью расходования
энергетических запасов организма во
время выполнения физических упражнений.
При правильном чередовании нагрузок и
отдыха в определенный период времени
после работы запасы энергетических
резервов превышают свой исходный (до
рабочий) уровень. Это явление получило
название сверхвосстановление (или
суперкомпенсация, см. рис. 2).
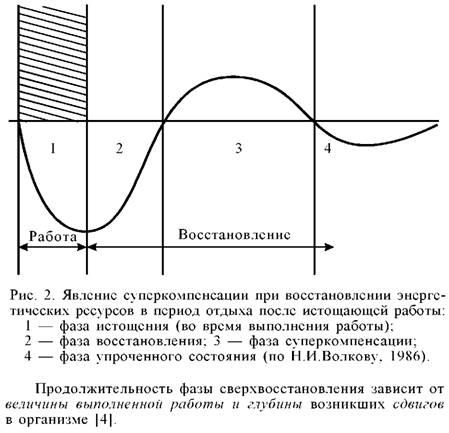
1.
После мощной, но кратковременной работы
эта фаза быстро наступает и быстро
завершается (например, восстановление
израсходованных запасов гликогена
отмечается через 34 часа, завершается
через 1 2 часов отдыха).
2.
После продолжительной работы умеренной
мощности восстановление гликогена
обнаруживается только через 1 2 часов,
а продолжительность фазы суперкомпенсации
продолжается в течение 4872 часов после
ее окончания.
Синтез
структурных белков в восстановительном
периоде происходит со скоростью более
низкой, чем скорость восстановления
энергоресурсов (в 46 раз). Поэтому после
напряженной работы, главным образом,
силового характера необходимо увеличить
продолжительность восстановления.
3.
В том случае, если несколько однонаправленных
нагрузок следуют одна за другой в фазе
не до восстановления, то после каждой
нагрузки степень недовосстановления
увеличивается. После уменьшения нагрузки,
изменяя ее направленность, или после
отдыха фаза суперкомпенсации выражена
более значительно и более продолжительна
во времени.
Ускорению
восстановительных процессов способствуют
также такие педагогические средства и
методы, как разнообразие условий
тренировки, создание благоприятного
эмоционального фона, оптимальные
биоклиматические условия, выполнение
дыхательных упражнений и др.
Психологические
средства восстановления физической
работоспособности спортсменов. С помощью
психологических средств снимают
состояние угнетенности, ускоряют
восстановительные процессы, снижают
уровень нервно-психической напряженности
и т.п. Для восстановления работоспособности
используются аутогенная тренировка
(включающая релаксацию и самовнушенный
сон), разнообразные формы досуга,
комфортабельные условия быта и другие
средства и методы восстановления
работоспособности.
Аутогенная
тренировка (от греч. autogenes — сам,
производящий)— это метод
психотерапии. В спорте при помощи
самовнушения спортсмен обучается
воздействовать на свое физическое и
психическое состояние. Основной задачей
в аутотренинге является достижение
приторможенного состояния коры больших
полушарий за счет снижения интенсивности
внешних и внутренних раздражителей.
Мысленно проговаривая необходимые
словесные формулы и вызывая в своих
мышцах ощущение тяжести, тепла и
расслабления, спортсмен самостоятельно
вводит себя в состояние полу- гипнотического
сна, или аутогенного погружения (см.
рис. 3).
Время
тренировки Рис. 3.Динамика процесса
аутогенной тренировки: 1 — начало
аутогенной тренировки; 2 — начало
аутогенного погружения; 3 — достижение
аутогенного погружения; 4 — начало сна;
5 — сон; 6 — активизация.
При
помощи аутогенной тренировки снижаются
физическая усталость, нервное напряжение,
нормализуется сон, спортсмен настраивается
на предстоящую деятельность, а также
лечатся некоторые заболевания.
Релаксация
(от лат. relaxatio— ослабление)— это
расслабление или резкое снижение тонуса
скелетной мускулатуры вплоть до полного
обездвижения.
Умение
спортсмена расслаблять мышцы во время
выполнения заданий, а также между ними
и после окончания тренировки и соревнований
способствует ускорению восстановительных
процессов. Эта способность тренируема.
Эффект повышается при расслаблении
мышц во время выполнения упражнений на
гибкость и дыхательных упражнений.
Релаксация
может достигаться и путем применения
миорелаксантов — лекарственных веществ,
вызывающих расслабление поперечнополосатых
мышц, которые, действуя на ЦНС, угнетают
структурные центры нервной системы,
регулирующие тонус этих мышц.
В
спортивной практике получила широкое
распространение одна из модификаций
аутогенной тренировки — психомышечная
тренировка (ПМТ). Она включает элементы
релаксации в сочетании с дыхательными
упражнениями и самовнушением. Основу
ПМТ составляют: а) умение максимально
расслаблять мышцы; б) способность, не
напрягаясь психически, но с предельным
воображением и как можно ярче, представить
содержание формул самовнушения; в)
умение концентрировать свое внимание
на какой-либо части тела; г) умение
воздействовать на самого себя мысленным
повторением словесных формул.
Действенным
средством воздействия на психику
человека является музыка. Она помогает
увеличить скорость восстановления
организма, повысить объем выполняемой
работы. Спокойная музыка делает дыхание
глубоким и ритмичным, быстрая дыхание
учащает. В периоды отдыха музыка должна
вызывать успокоенность, радость,
бодрость.
Усилить
воздействие музыки на психику человека
помогает цветомузыка: красный цвет
возбуждает; желтый ассоциируется с
чувством тепла, света, вызывает надежду;
зеленый и фиолетовый цвета успокаивают
человека.
Рациональное
питание как фактор восстановления
работоспособности спортсменов. Питание
спортсмена является важнейшим фактором
ускорения восстановительных процессов,
сохранения здоровья, повышения
работоспособности, достижения высоких
спортивных результатов. В практике
спорта оно имеет специфические
особенности, обусловленные повышенными
запросами организма, направленностью
тренировочных нагрузок, условиями
проведения соревнований и др.
Питательные
вещества выполняют энергетическую и
питательную функции.
Энергетическая
функция питания. На выполнение
тренировочных и соревновательных
нагрузок организм тратит энергию,
которая должна быть восполнена во время
выполнения упражнений (к примеру, питание
в марафонском беге) или в восстановительном
периоде. В противном случае спортсмен
не сможет выполнить последующие нагрузки
с достаточной эффективностью, невосполнение
энергетических запасов оказывает
отрицательное влияние на состояние его
здоровья (кроме случая, когда спортсмен
имеет «лишний» вес).
Энергия
в пище содержится в виде белков, жиров
и углеводов. В организме все они
расщепляются, образуя соединения, менее
богатые энергией. Высвободившаяся
энергия используется для нужд организма.
Об
энергетических запасах в пище судят по
ее калорийности.
Калория
(от лат. calor— тепло) — внесистемная
единица количества теплоты, обозначается
кал (1 кал = 4,1868 Дж).
С
энергетической точки зрения питательные
вещества могут взаимозаменяться в
соответствии с их калорической ценностью
(см. табл. 3).
Таблица
3.
Калорическая ценность питательных
веществ.
|
Питательные |
Жиры |
Белки |
Углеводы |
|
ккал/г |
9.3 |
4.1 |
4.1 |
|
кДж/г |
37.0 |
17.0 |
17.0 |
В
различных видах спорта спортсмены
тратят различное количество калорий
(см. табл. 4).
Таблица
4.
Состав
и калорийность пищевого рациона для
представителей различных видов спорта
(на 1 кг массы тела).
|
Вид |
Белки |
Жиры |
Углеводы |
Калорийность |
|
Гимнастика |
2,22,5 |
1,71,89 |
8,69,75 |
5966 |
|
Спринт, |
2,32,5 |
1,82,0 |
9,09,8 |
6267 |
|
Бег |
2,42,8 |
2,02,1 |
10,312,0 |
6978 |
|
Бег |
2,52,9 |
2,02,2 |
11,21 |
7384 |
|
Плавание |
2,32,5 |
2,22,4 |
9,510,0 |
6772 |
|
Тяжелая |
2,52,9 |
1,82,0 |
10,111,8 |
6677 |
|
Борьба |
2,42,8 |
1,82,2 |
9,011,0 |
6275 |
|
Гребля |
2,52,7 |
2,02,3 |
10,511,3 |
70,77 |
|
Футбол, |
2,42,6 |
2,02,2 |
9,610,4 |
6672 |
|
Баскетбол, |
2,32,4 |
1,82,0 |
9,510,8 |
6371 |
|
Велоспорт: |
2,32,5 |
1,82,0 |
10,811,8 |
6975 |
|
Велоспорт: |
2,52,7 |
2,02,2 |
12,214,3 |
7787 |
|
Конный |
2,12,3 |
1,71,9 |
8,910,0 |
6065 |
|
Парусный |
2,22,4 |
2,12,2 |
8,59,7 |
6268 |
|
Стрелковый |
2,22,4 |
2,02,1 |
8,39,5 |
6067 |
|
Лыжные |
2,32,5 |
1,92,2 |
10,211,0 |
6774 |
|
Лыжные |
2,42,6 |
2,02,4 |
11,512,6 |
7482 |
|
Конькобежный |
2,52,7 |
2,02,3 |
10,010,9 |
6974 |
При
составлении рациона питания необходимо
учитывать содержание в продуктах
питательных веществ (см. табл. 5).
О
полноценности пищи в энергетическом
отношении можно судить и по изменению
массы тела спортсмена. При достаточной
калорийности пищи вес спортсмена
практически не меняется. Увеличение
его в результате отложения жира указывает
на чрезмерное питание, а уменьшение —
на недостаточное.
Пластическая
функция питания.
В организме человека идет процесс
постоянного распада, превращения либо
обновления почти всех тканей. Для этого
в организм должно поступать минимальное
количество питательных веществ, другая
их часть может быть синтезирована самим
организмом.
Таблица
5.
Содержание
воды и питательных веществ в некоторых
пищевых продуктах (в весовых процентах)
|
Пищевые |
КДж |
Вода % |
Белки % |
Жиры % |
Углеводы |
|
Фрукты |
250 |
80,0 |
0,7 |
0,3 |
15,0 |
|
Овощи |
170 |
85,0 |
205 |
0,3 |
8,0 |
|
Картофель |
290 |
80,0 |
2,1 |
0,1 |
17,0 |
|
Сухой |
2400 |
1,8 |
5,3 |
40,0 |
50,0 |
|
Орехи |
2650 |
4,5 |
15,0 |
60,0 |
18,0 |
|
Хлеб |
1050 |
35,0 |
8,0 |
1,0 |
50,0 |
|
Мясо |
750 |
70,0 |
18,0 |
10,0 |
0,1 |
|
Колбаса |
1130 |
60,0 |
12,0 |
25 |
0,0 |
|
Масло |
3000 |
17,0 |
0,6 |
81,0 |
0,7 |
|
Сыр |
1400 |
45,0 |
23,0 |
27,0 |
3,0 |
|
Молоко |
270 |
89,0 |
3,1 |
3,4 |
4,65 |
|
Фруктовые |
170 |
85,0 |
0,3 |
0,1 |
12,0 |
Важное
место в рационе питания спортсмена
отводится белкам, т.к. во время мышечной
работы они интенсивно распадаются в
тканях. Белки состоят из отдельных
аминокислот, которые используются
организмом при обновлении и построении
тканей. Известно 24 вида различных
аминокислот. В состав пищи обязательно
должны входить белки, содержащие
незаменимые аминокислоты, т.к. они либо
совсем не образуются в организме, либо
образуются недостаточно. Белки повышают
также возбудимость нервной системы.
Показателем
биологической активности белка,
содержащегося в пище, может служить
количество белка организма, восполняющегося
при потреблении 1 00 граммов пищевого
белка. Для животного белка этот показатель
составляет 801 00 г (т.е. 1 00 г животного
белка может превратиться в 801 00 г белка
организма), а для растительных белков
— лишь 6070 г. Это связано с тем, что в
растительных белках содержание
незаменимых аминокислот находится в
неадекватном для человека соотношении.
Важную
роль в построении клеточных структур
(в частности, митохондрий — клеточных
«электростанций», осуществляющих
аэробный метаболизм) играют жиры.
Различают насыщенные и ненасыщенные
жирные кислоты. Ряд ненасыщенных жирных
кислот (к примеру, липоевая), необходимых
для жизнедеятельности, в организме не
синтезируется. После всасывания жиры
либо претерпевают окислительный распад
в процессе энергообеспечения организма,
либо откладываются в тканях, образуя
запас энергии.
Главным
источником энергии при выполнении
мышечной работы являются углеводы.
Энергетические потребности головного
мозга покрываются почти исключительно
за счет глюкозы. Скелетные мышцы,
напротив, при недостаточном потреблении
глюкозы расщепляют жирные кислоты.
Глюкоза используется также в качестве
строительного материала для синтеза
многих важных веществ организма.
Основными
углеводными молекулами являются простые
сахара — моносахариды. Соединения из
двух и более моносахаридов называются
ди, олиго или полисахаридами. Главным
углеводом в рационе питания человека
является крахмал. В организме углеводы
запасаются в виде гликогена — животного
крахмала.
Медико-биологические
средства восстановления физической
работоспособности.
В практике спорта нашли применение
физические, гидротерапевтические,
фармакологические и другие средства.
Рассмотрим основные из них.
Физические
средства восстановления работоспособности.
Они используются с профилактической и
оздоровительной целями для поддержания
высокой работоспособности и ускорения
восстановления, предупреждения
перетренированности, перенапряжений
и травм, а также при появлении начальных
признаков патологических процессов в
организме для ослабления их развития
и дальнейшего лечения . К этой группе
средств относятся ультрафиолетовое
излучение, аэроионизация, холодовые и
тепловые процедуры и др. Воздействуют
они через кожу. Физическое раздражение
рецепторов кожи оказывает рефлекторное
воздействие и на деятельность мышечной
системы, внутренних органов и ЦНС.
Гидротерапевтические
средства.
Наиболее известны из них дождевой,
холодный, горячий, контрастный душ,
различные ванны(пресная, горячая, ножная,
хлориднонатриевая, щелочная, хвойная
и др.). Гидротерапия способствует
регуляции кровоснабжения тканей и
ускорению окислительновосстановительных
процессов, выведению из организма
продуктов обмена, ликвидации застойных
явлений и микротравматических повреждений
в опорнодвигательном аппарате и др.
Фармакологические
средства восстановления работоспособности.
Биологически активные препараты находят
все более широкое применение в спортивной
практике. Они используются для ускорения
восстановления, активного восполнения
израсходованных пластических и
энергетических резервов, избирательного
управления функциональными системами
организма. Эти средства условно делят
на несколько групп.
Витаминные
препараты. В рационе питания спортсмена
обязательно должны присутствовать
витамины.
БИЛЕТ 31
(КРОВЬ КАК БИОЛОГИЧЕСКАЯ СИСТЕМА.ОСНОВНЫЕ
ФУНКЦИИ КРОВИ)
Кровь —
внутренняя среда организма, образованная
жидкой соединительной тканью.
Состоит из плазмы и
форменных элементов: клеток лейкоцитов и
постклеточных структур
(эритроцитов и тромбоцитов).
Циркулирует по системе сосудов под
действием силы ритмически
сокращающегося сердца и
не сообщается непосредственно с другими
тканями телаввиду
наличия гистогематических
барьеров.
В среднем, массовая доля крови к общей
массе тела человека составляет 6,5-7 %.
У позвоночных кровь имеет красный
цвет (от
бледно- до тёмно-красного), который ей
придаёт гемоглобин,
содержащийся в эритроцитах.
У некоторых моллюсков и
членистоногих кровь имеет голубой цвет
за счёт наличия гемоцианина.
Кровь
Понятие
о внутренней среде организма. Кровь,
лимфа и межтканевая жидкость являются
внутренней средой организма. Они
доставляют клеткам вещества необходимые
для жизнедеятельности, и уносят конечные
продукты обмена.
В
отличие от непрерывно изменяющейся
внешней среды внутренняя среда постоянна
по своему составу и физико-химическим
свойствам (температура, осмотическое
давление, реакция и др.). Постоянство
внутренней среды организма (гомеостаз),
по выражению великого французского
физиолога Клода Бернара, является
необходимым условием свободной жизни.
Например, высокоорганизованные
теплокровные животные могут жить при
температуре от + 50° С до — 72° С. Постоянство
температуры внутренней среды обеспечивает
постоянство жизнедеятельности всех
органов и систем организма.
Кровь
— это жидкая ткань, состоящая из плазмы
и взвешенных в ней кровяных телец.
Циркуляция крови по замкнутой
сердечно-сосудистой системе является
необходимым условием поддержания
постоянства ее состава. Остановка сердца
и прекращение движения крови немедленно
приводят организм к гибели.
Постоянство
состава и свойств крови регулируется
центральной нервной системой и железами
внутренней секреции.
Основные
функции крови
Находясь
в непрерывной циркуляции, кровь выполняет
транспортные функции:
1)
разносит по организму питательные
вещества;
2)
уносит от органов продукты распада и
доставляет их к органам выделения;
3)
участвует в газообмене, транспортирует
кислород (О2) и углекислый газ (СО2);
4)
поддерживает постоянство температуры
тела: нагреваясь в органах с высоким
обменом веществ (мышцы, печень), кровь
переносит тепло к другим органам и коже,
через которую происходит теплоотдача;
5)
переносит гормоны, метаболиты (продукты
обмена веществ) и осуществляет химическое
взаимодействие в организме, или
гуморальную регуляцию функций.
Кровь
выполняет защитную функцию. Она играет
главную роль в иммунитете — невосприимчивости
к инфекционным болезням и защите от
вредных агентов. К защитным функциям
крови относится также ее способность
к свертыванию, прекращающему кровотечение.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Описан креатинфосфатный путь ресинтеза АТФ в мышечных волокнах (креатинфосфатная реакция). Дана характеристика количественным критериям этой реакции: максимальной мощности, времени развертывания, времени работы с максимальной мощностью. Указаны виды спорта, в которых этот путь ресинтеза АТФ в мышечных волокнах является основным механизмом энергообеспечения.
Креатинфосфатный путь ресинтеза АТФ в мышечных волокнах
Определение
Креатинфосфатный (креатинкиназный ресинтез АТФ, алактатный ресинтез АТФ, креатинфосфатная реакция) – самый быстрый способ получения АТФ в мышечных волокнах.
Формула креатинфосфатной реакции следующая:
КрФ + АДФ → Кр + АТФ.
где: КрФ – креатинфосфат, Кр – креатин.
Расщепление КрФ в мышечных волокнах катализируется ферментом креатинкиназой. Активность этого фермента значительно возрастает при физических нагрузках за счет активирующего действия на неё ионов кальция, которые выделяются в саркоплазму из саркоплазматического ретикулума при мышечном сокращении.
Основные запасы КрФ организма человека находятся в мышечных волокнах. Доказано, что в мышечных волокнах II типа содержится на 10-15% больше КрФ, чем в мышечных волокнах I типа (В.Н. Платонов, 2005). В связи с этим считается, что в мышечных волокнах II типа креатинфосфатная реакция является основным путем ресинтеза АТФ.
Количественные критерии креатинфосфатного пути ресинтеза АТФ
Максимальная мощность
Максимальная мощность 900-1100 кал/мин кг. Это в три раза больше, аналогичного показателя для аэробного ресинтеза АТФ. Такая большая мощность данного пути ресинтеза АТФ определяется высокой активностью фермента креатинкиназы и высокой скоростью протекания данной реакции.
Время развертывания
Время развертывания 1-2 с. Столь малое время развертывания объясняется высокой скоростью протекания данной реакции.
Время работы с максимальной мощностью
Время работы с максимальной мощностью составляет 8-10 с. Это связано с малыми запасами КрФ в мышечных волокнах. В покое содержание КрФ в мышечных волокнах составляет 15-20 ммоль/кг. К концу 15-й секунды запасы КрФ уменьшаются вдвое. Через 45 секунд интенсивной физической работы запасы КрФ в мышечных волокнах подходят к концу.
В связи с тем, что креатинфосфатная реакция активно протекает в мышечных волокнах II типа, имеются неинвазивные тесты, позволяющие оценивать процент мышечных волокон II типа в скелетных мышцах (А.В. Шишкина, 2008; А.В. Самсонова с соавт., 2014).
Виды спорта, в которых креатинфосфатная реакция является основным механизмом энергообеспечения.
Столь мощное протекание данной реакции обусловливает виды спорта, в которых она является основным механизмом энергообеспечения: спринт (100 м), прыжки в высоту и длину, метание молота и диска, толкание ядра, толчок и рывок штанги; приседание со штангой, жим штанги лежа, тяга штанги и т.д.
Количество КрФ в мышечных волокнах возрастает на 70% при систематических тренировках, направленных на развитие скоростно-силовых качеств (Н.И. Волков с соавт., 2000).
Экономичность
Характеризуется низкой экономичностью.
Синтез КрФ в мышечных волокнах
Синтез КрФ в мышечных волокнах происходит во время отдыха путем взаимодействия креатина с избытком АТФ.
Кр + АТФ → КрФ +АДФ.
Литература:
- Волков Н.И., Несен Э.Н., Осипенко А.А., Корсун С.Н. Биохимия мышечной деятельности.- Киев: Олимпийская литература, 2000.- 504 с.
- Михайлов С.С. Спортивная биохимия. – М.: Советский спорт, 2009.– 348 с.
- Самсонова, А.В. Методы оценки композиции мышечных волокон в скелетных мышцах человека /А.В.Самсонова, И.Э.Барникова, М.А.Борисевич, А.В.Вахнин //Труды кафедры биомеханики НГУ им. П.Ф.Лесгафта.- вып. 6.- СПб, 2012.- С. 18-27.
- Шишкина А.В. Биодинамическая оценка мышечной композиции // Ученые записки университета имени П.Ф. Лесгафта, 2008. – №11. – С. 108-111.
С уважением, А.В. Самсонова
Похожие записи:
Сила тяжести
Дано определение силы тяжести. Показано, что сила тяжести является частным случаем силы гравитации. Описаны факторы, определяющие силу тяжести:…
Сила
Дано определение силы в механике. Описаны факторы, определяющие действие на тело силы: направление, точка приложения и численное значение.
Звенья тела человека как рычаги
Дано описание опорно-двигательного аппарата (ОДА) человека как системы рычагов. Приведен пример расчета силы двуглавой мышцы плеча…
Метаболический стресс. Накопление лактата в мышцах
Описан механизм влияния метаболического стресса (накопления лактата) на гипертрофию мышечных волокон. Показано, что накопление лактата приводит к…
Механическое повреждение мышечных волокон
Описаны механизмы механического повреждения мышечных волокон при силовой тренировке, приводящие к гипертрофии скелетных мышц. Показано, что…
Механическое напряжение (механотрансдукция) в скелетных мышцах
Описаны процессы передачи механического напряжения в скелетных мышцах. Показано, что механическое напряжение, возникающее вследствие сокращения скелетных…
Двигательная активность: утомление и восстановление

- В далёкие времена человеку для своего
жизнеобеспечения нужно было быть сильным, быстрым, ловким. Только так он мог
добывать пищу и защищаться. В естественных природных
условиях человек существовал тысячи лет. В результате угрозы возникали
стрессовые ситуации, которым всегда сопутствовала последующая физическая
нагрузка в виде бега или борьбы. Стрессовые ситуации мобилизировали организм для
преодолению предстоящей нагрузки. Возбуждение нервной системы происходило
естественно в рамках физиологической и биологической разрядки и не имело
никаких отрицательных последствий для организма. - Хочется отметить, что и пища человека
была простой, религиозные обряды и посты выполнялись неукоснительно, а если задуматься,
то это является благом. Что касается движения, оно всегда было естественной
необходимостью для обеспечения жизни. - В настоящее время стрессовые ситуации
не имеют разрядки в виде физической нагрузки, в связи с чем накапливаются
огромные отрицательные последствия для здоровья человека, которые
сопровождаются повышением артериального давления, головными болями, а в
конечном итоге – болезнями. - Как утверждает известный врач
кинезитерппевт профессор С. Бубновский, …боли возникают не в костях, как
кажется многим, а в мышцах, связках, сухожилиях. Питательные вещества для
костей поступают через мышцы, а их обмен происходит только в тех мышцах,
которые работают. Если мышцы не работают, то в костях возникают дистрофические
изменения». Принято считать, что это возрастное явление. По утверждению Бубновского,
всё не так. Как только мышцы включаются в работу, проблемы исчезают. Вот почему
человеку нужны постоянные существенные физические нагрузки, которые позволяют
защитить его от стрессовых ситуаций и сохранить необходимый уровень здоровья. - Движение всегда являлось
катализатором продолжительной и здоровой жизни. Известны слова французского
врача Тиссо: «Движение как таковое может заменить любое лекарство, но все
лечебные средства мира не в состоянии заменить действие движения». Сегодня мы
можем привести множество примеров, подтверждающих эти слова, начиная от
Валентина Дикуля, известного циркового артиста, получившего тяжелейшую травму,
когда сила воли и физические упражнения полностью восстановили его физическую
работоспособность. - Директор научно-исследовательского
центра сердечно-сосудистой хирургии им. Бакулева академик Лео Бокерия, внесённый
в список 100 лучших хирургов мира, в одном из интервью на вопрос, как
приумножить здоровье, отметил, что определяющее значение имеет то, что заложено
в детстве. Родители до 10–12 лет должны успеть привить ребёнку тягу к движению,
спорту. Именно в этот период вырабатывается трудолюбие, выносливость,
координация, что очень важно для работы сердца. Если человека не приучить к
этому в детстве, во взрослой жизни заставить его следить за собой и своим
здоровьем практически невозможно. По словам Бокерия, специалисты знаменитой
кардиологической клиники братьев Мейо в США рекомендуют взрослым людям, которые
никогда не занимались спортом, начинать с одного часа активной ходьбы в неделю.
Через месяц продолжительность прогулок можно увеличить до двух часов. Вот так
потихоньку у человека возникает потребность в физической нагрузке. - Исследованиями доказано, что
двигательная деятельность является физиологической потребностью человека и всех
живых организмов, она не менее важна, чем питание, сон, утоление жажды.
Недостаточность или отсутствие двигательной активности отрицательно сказывается
на формировании молодого организма, нормальной жизнедеятельности взрослого
человека: развиваются различного рода заболевания, обнаруживается
недостаточность обмена веществ, ускоряются и усиливаются процессы увядания и
старения. Эти изменения особенно активизируются, если недостаточная физическая
активность дополняется систематическими эмоциональными напряжениями и
перенапряжениями. - Современная жизнь, связанная с
автоматизацией и компьютеризацией условий труда и быта, создала условия для
малоподвижного режима профессиональной деятельности и отдыха. Такой режим
называется гиподинамией. Она характеризуется снижением тонуса мускулатуры,
возникновением неблагоприятных физиологических процессов в организме человека,
недостаточным количеством сигналов, поступающих в центральную нервную систему
от рецепторов опорно-двигательного аппарата, нарушением баланса в работе
нервных центров и внутренних органов, снижением потенциальных возможностей человека
и, прежде всего, сердечно-сосудистой системы и физических качеств. - Регулярное выполнение физических
упражнений с широкой амплитудой движений оказывает механическое воздействие на
различные сосуды, проходящие через позвоночник, путём сдавливания, растягивания,
массирования. Многократное их выполнение способствует сохранению эластичности
стенок сосудов, увеличению их прочности, препятствует склеротическим изменениям.
Что же касается воздействия физических упражнений на состояние позвоночника,
оно заключается в улучшении его гибкости, подвижности, растяжимости,
стабильного состояния. Следует применять также комбинированные упражнения,
которые более динамичны, эффективны, эмоциональны и их с удовольствием
выполняют занимающиеся. - Регулярные физические нагрузки
укрепляют и омолаживают организм, создают необходимые резервные возможности. Можно
заниматься разными физическими упражнениями: атлетической или ритмической
гимнастикой, спортивными играми, циклическими видами спорта. Одни упражнения
развивают силовые качества, другие – скоростно-силовые, третьи – общую и
специальную выносливость. Японские (и не только) учёные полагают, что
незаменимым средством оздоровления является ходьба. 1000 шагов в день – минимальная
доза физической активности, необходимая организму человека для его оптимального
жизнеобеспечения. - Для того чтобы укрепить здоровье,
повысить работоспособность, увеличить свой физиологический резерв, необходимы
систематические занятия различными физическими упражнениями или видами спорта.
Установлено, что с возрастом физиологический резерв организма уменьшается. Уже
в 30–40 лет у лиц, ведущих малоподвижный образ жизни, не занимающихся
физическими упражнениями или другими видами физического труда, могут появиться
признаки атеросклероза, уменьшается жизненная ёмкость лёгких, снижается
мышечная сила, ухудшается координация движений, подвижность суставов,
эластичность мышц, происходят другие неблагоприятные изменения. - Каждый человек должен примерно знать
и оценивать «крепость» своего здоровья: какие у него кровяное давление, частота
пульса, содержание сахара и гемоглобина в крови, степень одышки при нагрузке,
общее состояние желудочно-кишечного тракта, сердечно-сосудистой и нервной
систем. Тренировка резервов организма должна быть постепенной, но настойчивой,
имеется в виду увеличение количества движений, скорости и расстояния. - Специалисты в области физиологии,
спортивной медицины, педагогики призывают нас учиться у природы. Посмотрите, как
работает сердце: сокращение – расслабление, активность – покой. Такой
рациональный режим изобрела природа. Поэтому человек, не придерживающийся
режима, подсказанного природой, быстро утомляется, теряет активность и интерес
к работе, часто болеет. Избежать подобных явлений можно: переключаясь с одного
вида деятельности на другой, вы невольно будете менять положение своего тела,
включая в работу другие органы, когда кровь будет перераспределяться в
организме, а снабжение ею в различных органах будет не одинаково. - При любой мышечной деятельность
возникает утомление, характеризующееся комплексом изменений состояний различных
функций организма. Чем больше степень выраженности этих изменений, в том числе
чувства усталости, тем интенсивнее и продолжительнее была выполненная работа. - Таким образом, утомление – это особое
состояние, возникающее как следствие работы и проявляющееся в ухудшении
двигательных и вегетативных функций, понижении работоспособности и появлении
чувства усталости. Это состояние имеет временный характер и исчезает через
некоторое время после прекращения работы, т. е. во время отдыха до периода полного
восстановления. Усталость – это субъективное проявление утомления, которое
выражается в определённом эмоциональном состоянии, ощущении тяжести в
работающих мышцах или группе мышц, появлении желания прекратить работу или
уменьшить физическую нагрузку. - Усталость отражает физиологическое
состояние систем организма человека, обеспечивающих его работоспособность и
особенности физического развития, расходования и восстановления активных
энергетических веществ. При интенсивной и продолжительной работе может
наступить момент, когда возникает чувство усталости, усиливаемое отрицательными
эмоциями, а часто и ощущение тяжести и даже боли в работающей системе
организма. Степени усталости и утомления могут не совпадать по времени, что
зависит от положительного или отрицательного эмоционального фона, на котором
осуществляется физическая активность. Устать можно и от безделья. И наоборот:
увлечённый работой человек не ощущает усталости даже тогда, когда некоторые
физиологические сдвиги свидетельствуют о наступившем утомлении. - Выделяют различные признаки утомления: физиологические,
психологические, медицинские и другие. С физиологической стороны развитие
утомления свидетельствует о значительном сокращении внутренних резервов
организма человека на менее выгодные виды режима работы. Вследствие этого
изменяется темп работы, нарушается точность, ритмичность, координация движений,
техника их выполнения, изменяется характер протекания психических процессов. В
зависимости от конкретных условий двигательной активности на первый план в
развитии утомления выходят различные факторы. Это следует понимать так, что
физиологические проявления утомления зависят от характера его возникновения.
Каждый вид работы (физической, умственной), связанной с напряжением внимания
или процессами мышления приводит к появлению различных форм утомления. - С биологической точки зрения
утомление играет определённую роль. Во-первых, ощущение усталости выполняет
защитную функцию, представляя физиологическую меру против истощения. Во-вторых,
процесс расходования функциональных ресурсов стимулирует интенсивность
восстановительных процессов и именно этим в итоге определяется постепенное
нарастание функционального потенциала под влиянием упражнений и тренировок.
Повторное и не чрезмерное утомление является действенным фактором повышения
функциональных возможностей организма и его работоспособности. - В целом, при физической и умственной
работе утомление проявляется тем больше, чем меньше физиологические резервы
организма. Поэтому у детей утомление может развиваться быстрее, чем у взрослых.
При высоких нагрузках (физической и умственной деятельности) без достаточного
отдыха для восстановления утомление может перейти в хроническую форму и вызвать
ухудшение работоспособности и самочувствия. В этом случае важно организовать
свою активность так, чтобы умственные нагрузки разумно чередовались с
физическими, то есть чтобы смена деятельности стала формой активного отдыха. - Специфика физиологических процессов
при различных видах двигательной активности обуславливает различную природу
возникновения и протекания утомления. Существенное значение при этом имеет
характер деятельности, её длительность, мощность, сложность упражнений и др.
Например, при выполнении циклической работы умеренной мощности (бег,
передвижение на лыжах, велосипеде, плавание с частотой пульса 130–140 ударов в
минуту) основной причиной развития утомления является трудность длительного
поддержания на необходимом уровне физиологических функций. При работе такой
мощности, продолжительностью более 40–60 минут, уменьшается содержание сахара в
крови, что может негативно повлиять на деятельность центральной нервной системы,
а значительная потеря ионов натрия, калия и кальция, хлора и фосфора в крови и
тканях тела ведут к понижению работоспособности и усталости. - Появлению утомления способствует
перегревание организма, которое возникает при длительной умеренной работе в
условиях высокой температуры и влажности окружающей среды. - Следует отметить, что утомление в
процессе мышечной или умственной деятельности, не превышающее допустимых
пределов, – физиологическое, а не патологическое явление, полезно для организма
человека, так как представляет собой важный фактор роста подготовленности
(тренированности), когда оно связано с развитием выносливости, необходимой
каждому человеку. Физиологическая особенность этого явления заключается в том,
что, тренируясь до наступления утомления, человек адаптируется к более высокому
уровню работоспособности. В тех же случаях, когда физическая нагрузка
прекращается до начала возникновения утомления, повышение работоспособности
организма приостанавливается. То же происходит, если физические нагрузки приводят
к резко выраженному утомлению. В связи с этим при выполнении физических
нагрузок спортивной или профессиональной деятельности следует избегать не
утомления вообще, а лишь чрезмерного его уровня. - Важным средством борьбы с утомлением
и переутомлением является рациональный режим труда и отдыха или организация в
установленное время кратковременных перерывов. Такие перерывы должны
устраиваться с учётом характера и длительности выполняемых двигательных или
умственных нагрузок. Известно, что работа, выполняемая ритмично, примерно на 20 %
менее утомительна. - Двигательная активность, как правило,
сопровождается временным снижением работоспособности. После окончания работы
наступает период восстановления, при котором нормализуется внутренняя среда
организма, восстанавливаются энергетические запасы, различные функции приходят
в состояние рабочей готовности. Эти процессы не только обеспечивают
восстановление работоспособности организма, но и способствуют её увеличению. - Восстановительные процессы начинают
протекать непосредственно во время мышечной работы. Примером этого являются
окислительные реакции, обеспечивающие ресинтез богатых энергией химических
веществ. Однако при работе процессы диссимиляции преобладают над процессами
ассимиляции. Лишь при длительной мышечной активности устанавливается равновесие
между расщеплением химических веществ и их ресинтезом. - В восстановительном периоде
преобладают процессы ассимиляции. Они обеспечивают пополнение израсходованных
при двигательной активности энергетических запасов. Сначала они
восстанавливаются до исходного уровня, а затем на некоторое время становятся
выше, то есть превышают исходный уровень (фаза сверхвосстановления). Если в это
время не предлагается нагрузка, работоспособность вновь понижается. Вот почему
необходимо соблюдать принципы и закономерности физиологических изменений в
организме человека. В связи с этим повторные нагрузки целесообразно выполнять в
фазу повышенной работоспособности. В этих условиях тренированность организма
развивается наиболее интенсивно. Если этого не происходит, то работоспособность
снижается до исходного уровня и в дальнейшем прогрессивные изменения в
организме приостанавливаются. - О восстановлении работоспособности
организма можно судить по восстановлению отдельных его функций, например по
динамике частоты сердечных сокращений. Этот показатель определяется сразу после
работы, а затем повторно, через определённые промежутки времени. Снижение
частоты сердечных сокращений, установленной после работы, позволяет судить об
интенсивности восстановления организма. - Выполнение длительной работы большой
и умеренной мощности характеризуется медленным восстановлением дыхательных
функций и энергетики. Даже у квалифицированных спортсменов энергетические траты
снижаются до исходных величин очень долго – до нескольких суток. Например, у
тренированных лыжников это продолжается 2–3 дня. - Изучение восстановительных процессов
при развитии физических качеств (быстроты, силы, выносливости) показывает, что их восстановление происходит по-разному
(и по времени, и по эффективности). Даже взаимосвязанные двигательные действия,
развивающие силу и силовую выносливость, восстанавливаются в разное время.
Повторность выполнения физических упражнений на занятиях должна планироваться с
учётом того, что эффективность последующей работы будет больше тогда, когда
утомление от предыдущей работы почти ликвидировано, а положительное воздействие
ещё сохранено. Чтобы процессы восстановления протекали в нужном направлении,
следует, как минимум, не мешать организму выполнять данную функцию. Например,
если сходить в спортзал, поплавать в бассейне или долгое время кататься на
велосипеде, то после таких двигательных действий необходимо хорошо поесть и
поспать. Если израсходованная энергия и питательные вещества не восполняются, а
продолжительность сна будет недостаточной, то отсутствие «свежести и бодрости»
на следующий день будет закономерным исходом, а для того, чтобы после
физических нагрузок организм не просто восстанавливался, а восстанавливался
быстро, необходимо выполнять определённые действия. - При активной мышечной работе
возникает потребность в употреблении продуктов с повышенной биологической
ценностью, то есть питательные вещества в наибольшей концентрации
(аминокислоты, жирные кислоты, витамины, соли и др.). Существует достаточное
количество безопасных веществ природного происхождения, которые помогают нам в
этом, – некоторые аминокислоты (триптофан, метионин, лезин, креатин) и другие витаминоподобные
вещества. - Особое внимание необходимо уделить
продуктам питания и калорийности пищи. Энергию из пищи целесообразнее извлекать
из каш, овощей, фруктов, ягод, корнеплодов и молочных продуктов. Но
непосредственно перед и сразу после физических нагрузок можно рекомендовать
фруктовые соки и лёгкие мучные изделия. Они имеют высокий гликемический индекс,
то есть быстро перевариваются и всасываются в кровь. - Сразу после физической нагрузки
возникает потребность в восполнении израсходованного гликогена и так как гликоген
образуется из глюкозы, то и первоочередной задачей является употребление
продуктов её содержащих. А когда в кровь поступает большое количество глюкозы,
поджелудочная железа выбрасывает в кровь инсулин, который способствует
быстрейшему восстановлению организма. - Необходимо помнить, что употребление
мучного и сладкого не должно стать привычкой. Наиболее ценными будут и другие
продукты, обладающие высоким гликемическим индексом, – мёд, финики, абрикосы,
морковь, кабачки, орехи. В отличие от мучных продуктов и сахара, они
биологически более ценные, так как содержат много полезных комплексов
(витамины, микроэлементы, органические кислоты и др.). - Восстановление запасов гликогена
является предпосылкой для начала механизма восстановления других биологических структур
– белков, а началом их обмена является окончание физической нагрузки. Но синтез
белка является довольно энергозатратным, своеобразным ответом на нагрузку. И на
этот ответ следует обращать внимание занимающимся физической активностью, а не
только профессионально тренирующимся спортсменам. Желательно, чтобы продукты
питания содержали и быстрые для усвоения белки: рыбу, орехи, яйца, молочную
сыворотку и др. - Любой интенсивный труд немыслим без
потери жидкости – то есть потения. Учитывая, что источником жидкости для пота
является плазма крови, становятся понятными такие последствия, как уменьшение
объёма циркулирующей крови: сгущение крови, нарушение солевого баланса. Чем это
грозит? Вода – среда, в которой протекают все биохимические реакции. Для
нормального их протекания не должно быть дефицита воды в биологических
жидкостях (крови, лимфе, жидкости в межклеточном пространстве). Всё это нужно
для того, чтобы к клеткам быстрее доставлялся кислород и питательные вещества,
быстрее выводились продукты обмена веществ, восстанавливались и обновлялись
ткани. Подумайте, как будет чувствовать себя человек, занимающийся любой
двигательной деятельностью, который по своей неопытности и незнанию не
побеспокоился о своевременном употреблении жидкости (воды). Во-первых, быстро
наступает утомление, во-вторых, работоспособность восстановится значительно
позже и будет происходить медленно. Пить воду следует не тогда, когда хочется,
а до появления жажды, ибо жажда – признак уже наступившего обезвоживания с
вытекающими указанными последствиями. - Практика показывает, что только
совокупность использования педагогических, медико-биологических,
психологических средств и методов может составить наиболее эффективную систему
восстановления организма человека после активной двигательной нагрузки. - Валентин Киселёв,
- кандидат
педагогических наук, профессор кафедры физического воспитания и спорта
Белорусского государственного университета; - Вячеслав Харук,
- старший
преподаватель кафедры физического воспитания и спорта Белорусского
государственного университета; - Марьян Мазуро,
- старший
преподаватель кафедры физического воспитания и спорта Белорусского
государственного университета