Научная статья Евгения Суборова, к.м.н., врача анестезиолога- реаниматолога о роли гемоглобина в жизни бегунов.
Мы продолжаем публикацию научных статей Евгения Суборова о физиологии бега. Вас ждет исчерпывающий рассказ о том, как меняется уровень гемоглобина во время бега, что такое «спортивная анемия» и как тренировки влияют на вязкость крови.
Если у вас останутся вопросы, или вы хотите узнать больше об этой теме, задавайте вопросы в комментариях к этой статье.
От автора
Существуют разные способы повышения уровня гемоглобина у спортсменов: одни из них — легитимны, другие — нет. Главное — не терять голову и помнить, что избыточно высокий уровень гемоглобина может нанести вред здоровью.
Для чего нужен гемоглобин?
Гемоглобин содержится внутри красных кровяных телец — эритроцитов, которые
отвечают за транспорт кислорода и углекислого газа. Во время вдоха в легкие попадает кислород, который доходит до дыхательных мешочков — альвеол, а дальше, через тончайшую мембрану он переносится в микрососуд (капилляр). В капилляре кислород попадает в эритроцит, содержащий гемоглобин, в результате образуется молекула гемоглобина, связанного с кислородом — оксигемоглобина.
Эритроциты доставляют оксигемоглобин в разные ткани организма (включая мышечную), там гемоглобин «разгружается», теряет кислород, превращаясь в дезоксигемоглобин
Отдав кислород, гемоглобин присоединяет углекислый газ, который образуется в клетках в результате процессов обмена, представляя собой те «отходы», которые необходимо вывести. Попадая в легкие, углекислый газ переходит из эритроцитов в альвеолы, выделяется в атмосферу, а освободившееся место занимает кислород.
Процесс обмена кислорода и углекислого газа в легких называется альвеолокапиллярная диффузия.
То, насколько важен транспорт кислорода, становится очевидно при развитии анемии, когда снижение концентрации гемоглобина может вести к снижению производительности (1, 2). Однако, важна не только концентрация, но и функциональная «полноценность» гемоглобина. Например, в условиях недостаточного содержания кислорода (высокогорье), повышается способность гемоглобина связывать кислород в лёгких, обеспечивая нас жизненно необходимым газом (3). Удовлетворить возросшие потребности в кислороде при физической работе позволяет способность гемоглобина легче отдавать кислород в работающей мышце (4). Эти примеры показывают гибкость молекулы гемоглобина, способность подстраиваться под условия окружающей среды и предупреждать развитие кислородного голодания.
Помимо транспорта кислорода, эритроциты выполняют и другие функции, влияющие на производительность спортсмена:
— Способствуют поддержанию постоянства внутренней среды организма.
— Могут поглощать ряд продуктов обмена, выделяющихся при интенсивной работе мышц, например, лактата (молочной кислоты).
— Способствуют поддержанию проходимости сосудов, выделяя оксид азота (вещество, расширяющие сосуды) (5), что важно при физической работе (6).
Давайте разберемся, с помощью каких механизмов эритроциты обеспечивают непрерывную доставку кислорода к тканям, позволяя выполнять физическую работу на протяжении длительного времени.
Почему так важна прочность связи гемоглобина с кислородом?
Изменение прочности связи гемоглобина с кислородом — один из основных механизмов, который оптимизирует транспорт кислорода, он не зависит от концентрации кислорода и общей массы гемоглобина.
К чему приводит изменение прочности связи гемоглобин-кислород?
Гемоглобин — это транспортный белок, выполняющий две противоположных задачи — присоединение кислорода в легких и отдача его тканям. Гемоглобин должен хорошо присоединять кислород, но при слишком сильной связи с гемоглобином, кислород будет неохотно отдаваться тканям на периферии.
Прикрепившись к гемоглобину, самостоятельно кислород от него отсоединиться не может (7). Для разрыва связи необходимо воздействие на гемоглобин одного из внешних факторов (например, pH, изменение концентрации углекислого газа, изменение температуры, а также изменение концентрации вещества со сложным названием 2,3-дифосфоглицерат). Изменение прочности связи гемоглобин-кислород может идти в двух направлениях: гемоглобин легче присоединяет кислород в легких (это имеет значение на высокогорье, где кислорода мало), или же легче отдает кислород тканям (например, при физической работе, когда мышцы требуют большого количества кислорода).
Увеличенная потребность в кислороде во время тренировок удовлетворяется за счет повышения кровотока в мышцах (8) и облегчения процесса высвобождения кислорода в тканях (9), тогда как лактат, накапливающийся в мышцах при физической нагрузке, практически не влияет на обеспечение тканей кислородом (10). Кроме этого, у тренированных спортсменов эритроциты образуются более активно, продолжительность их жизни уменьшается (11), более молодые эритроциты имеют повышенную метаболическую активность, а прочность связи гемоглобин-кислород в тканях ниже (12). Это означает, что у тренированных спортсменов кислород легче отдается тканям, что абсолютно необходимо при тяжелых и интенсивных тренировках.
Разгрузка кислорода в работающих мышцах
Работающие мышцы выделяют в кровеносные капилляры протоны водорода, углекислый газ и лактат, а температура в мышцах повышается до 41°C. Кровоток, проходящий через работающие мышцы, реагирует на эти изменения и активно отдает кислород мышцам (13), например, за счёт более высокого уровня 2,3-ДФГ в крови (14).
Обогащение крови кислородом в лёгких
На пути крови из мышцы в лёгкие происходит снижение концентрации H+, накопление углекислого газа, а также снижение температуры (температура крови в легких ниже, чем в работающих мышцах). Все это должно приводить к облегчению загрузки кислорода в эритроциты и гемоглобин, однако, на фоне интенсивных тренировок, кислород в легких поглощается не так активно, как в состоянии покоя. Это приводит к снижению максимального насыщения артериальной крови кислородом в покое с 97,5% до 95%. Другими словами, гемоглобин не полностью загружен кислородом, остается еще «свободное место». Компенсацией за эту неполную загрузку, является кислород, эффективно и активно высвобождающийся в работающих мышцах, что позволяет обеспечивать кислородом активно работающие мышцы (15).
Транспорт кислорода
В одном литре крови может растворяться только 0.03 мл кислорода, а грамм гемоглобина может переносить 1.34 мл O2. Таким образом, нормальное содержание гемоглобина в единице объема крови позволяет переносить достаточное количество кислорода для адекватного обеспечения тканей. Повышение уровня гемоглобина увеличивает количество доставляемого тканям кислорода. Способность переносить кислород влияет на производительность, которая повышается, например, после переливания эритроцитарной массы (16). Кроме того, описана зависимость между общим гемоглобином и максимальным потреблением кислорода (МПК или VO2max) у спортсменов (17). Следовательно, хорошая производительность спортсмена отчасти определяется высокой транспортной способностью крови.
Что влияет на способность крови переносить кислород?
Конечно, это концентрация гемоглобина в крови (cHb), гематокрит (Hct), общая масса гемоглобина (tHb), общий объем эритроцитов (tEV) в крови. Как cHb, так и Hct легко измерить при взятии образца крови на анализ. Вместе с показателем насыщения гемоглобина кислородом они показывают количество кислорода, которое может быть доставлено к тканям. Показатели tHb и tEV характеризуют общее количество кислорода, которое может транспортироваться кровью, высокие цифры этих показателей позволяют перераспределять кислород к органам и тканям с максимальной потребностью, поддерживая в то же время базовое поступление кислорода к менее активным органам и тканям. Примером активной ткани в беге может служить мышечная ткань — она требует много кислорода. А неактивная — это, например, ткань кишечника.
Гематокрит у спортсменов
Гематокрит, или объем красных кровяных клеток (эритроцитов) в крови, определяет способность крови переносить кислород. Большинство исследований показывают, что гематокрит спортсменов ниже, чем у нетренированных людей (18). Чрезмерно повышенный гематокрит увеличивает вязкость крови, что приводит к нарушениям работы сердечно-сосудистой системы (19).
Изменения гематокрита развиваются очень быстро, а выраженность изменений зависит от интенсивности и типа тренировок (20). Во время тренировок гематокрит может повышаться из-за уменьшения объема плазмы, особенно при недостаточном восполнении жидкостью (21). Низкий уровень гематокрита после тренировки объясняется быстрым увеличением объема жидкой части крови (плазма). Объем эритроцитов остается неизменным в течение нескольких дней (22), а «дотренировочные» уровни гематокрита восстанавливаются через несколько недель (24). Кроме того, уровень гематокрита подвержен и сезонным влияниям, летом он может быть ниже на 1-2%, в дополнение к снижению, вызванному тренировками (25).
Снижение уровня гематокрита у спортсменов называется «спортивная анемия». Долгое время это объяснялось повышенным разрушением эритроцитов во время тренировки, и, по сути, напоминает известный феномен под названием «маршевая гемоглобинурия». Ее также называют «болезнью солдат», поскольку механическое разрушение эритроцитов связано с чрезмерной нагрузкой на стопу. Первым признаком, которым проявляет себя гемоглобинурия, считается окрашивание мочи в темно-красный цвет, что объясняется присутствием в моче большого количества оксигемоглобина (26). У спортсменов внутрисосудистое разрушение эритроцитов связано с интенсивностью и типом тренировки, а ударная нагрузка на стопу является одной из самых частых причин, причем, она может быть частично предотвращена с помощью хорошо амортизирующей обуви (27). Другими возможными причинами «спортивной анемии» может быть недостаточное потребление белка, а также нарушенный липидный профиль и недостаток железа в организме (28).
Как было сказано выше, объем плазмы изменяется достаточно быстро, тогда как изменения общей массы эритроцитов происходят очень медленно, из-за невысокой скорости образования эритроцитов (29). Таким образом, измерение этих двух показателей, наряду с гемоглобином и гематокритом, помогает определить способность крови переносить кислород.
В ряде исследований было показано, что у тренированных спортсменов уровень tHb повышен (30), а повышение tHb на 1 г. увеличивает VO2max примерно на 3 мл/мин (31). Доказано, что повышение tHb на 1 г/кг массы тела повышает VO2max примерно на 5.8 мл/мин/кг, причем у нетренированных людей (даже у тех, кто имеет нетипично высокий показатель VO2max 45 мл/мин/кг) tHb = 11 г/кг, а у хорошо тренированные спортсмены (средний VO2max = 71.9 мл/кг) tHb = 14.8 г/кг (32).
Эти находки подтверждают данные 1949 года о том, что у элитных спортсменов tHb на 37% выше, чем у нетренированных людей (33). Однако, изучение tHb во время тренировочного процесса показало, что этот показатель изменяется очень медленно, и а выраженный рост возможен только после нескольких лет тренировок (34). Например, за период 9-месячного тренировочного цикла tHb увеличивается лишь на 6%.
Показатели tHb у жителей высокогорья выше по сравнению с жителями равнин (35), но для повышения tHb необходимо находиться в условиях высокогорья в течение нескольких недель или даже месяцев, тогда как кратковременное пребывание на высоте не повысит tHb и tEV (36). В одной работе повышение tEV было зафиксировано только после 3-х недельного пребывания в условиях высокогорья (37).
Влияние тренировок на образование эритроцитов
Повышение tHb и tEV у спортсменов доказывает, что тренировки стимулируют эритропоэз. Дополнительным признаком этого служит повышение уровня ретикулоцитов (клетки-предшественники эритроцитов в процессе кроветворения, составляющие около 1% от всех циркулирующих в крови эритроцитов), развивающееся через 1-2 дня после тренировки (38). Несмотря на очевидный эффект тренировок, в ряде исследований было показано, что количество ретикулоцитов у спортсменов не сильно отличается от нетренированных людей, а уровень этих клеток достаточно стабилен в течение многих лет (39). Вариабельность количества ретикулоцитов у спортсменов в течение года связана, как правило с интенсивным тренировочным процессом. В начале сезона количество ретикулоцитов максимально, а на фоне тяжелых тренировок, соревнований, а также в конце сезона их уровень снижается (40).
На эритропоэз влияет ряд факторов, которые изменяются под влиянием тренировок. Содержание мужских половых гормонов, временно повышающийся после тренировки, воздействует на эритропоэз путем стимуляции выработки ЭПО (эритропоэтин, один из гормонов почек, который контролирует образование красных кровяных клеток), что повышает активность костного мозга, включение железа в эритроциты, и проявляется резким повышением количества эритроцитов (полицитемия) (41, 42). Интересно, что уровень тестостерона после тренировки или соревнования изменяется в зависимости от настроения (выиграл/проиграл), причем этот эффект более выражен у мужчин (43).
Есть мнение, что стрессовые гормоны (адреналин, кортизол) стимулируют высвобождение ретикулоцитов из костного мозга и усиливают эритропоэз (44). Кроме того, эритропоэз стимулируется гормоном роста и инсулиноподобными факторами роста (45), которые также повышаются во время тренировок (46).
Вязкость крови
Гематокрит не только влияет на количество кислорода, которое может переносить единица объема крови, но изменяет и вязкость крови. Чем выше уровень гематокрита, тем выше вязкость и сопротивление току крови, что повышает нагрузку на сердце и приводит нарушениям кровотока. Частично компенсировать повышение вязкости при высоких цифрах гематокрита может способность эритроцитов изменять свою форму, что позволяет им проходить даже в очень небольшие по диаметру сосуды (47). Хорошо известна, например, пулеобразная форма (bullet-like shape) эритроцитов.
Тренировки активно влияют на вязкость крови. Во время тренировки повышается вязкость крови (48), одной из основных причин этого является недостаточный прием жидкости (49), а также нарушение способности эритроцитов изменять свою форму (50, 51, 52). Повышение лактата во время тренировок в целом не влияет на деформацию эритроцитов (53), но есть данные о том, что у тренированных спортсменов лактат может улучшить способность эритроцитов к деформации (54).
Собранные вместе данные говорят о том, что повышение вязкости крови во время тренировки вызвано повышением вязкости плазмы и снижением пластичности эритроцитов, что ухудшает доставку кислорода к работающим мышцам. Тем не менее, тренировки могут способствовать снижению вязкости крови (55, 56, 57), поскольку вырабатываются «защитные механизмы»: повышение выработки юных эритроцитов, выделение оксида азота, который способствует поддержанию проходимости сосудов (58). Все это позволяет поддерживать нормальную функцию сердечно-сосудистой системы и обеспечивать мышцы достаточным количеством кислорода.
Резюме
Существует множество механизмов, способствующих поддержанию нормальной доставки кислорода к работающим мышцам. Повышенные запросы в кислороде во время физической нагрузки обеспечиваются увеличением сердечного выброса и кровотока в мышцах, перераспределением кровотока (когда кровоток преимущественно уходит к органам и тканям, бесперебойная работа которых важна в беге), а также путем оптимизации кровотока в микрососудах, где происходит отдача кислорода (59). Эритроциты поддерживают проходимость сосудов и нормальный кровоток за счет выработки оксида азота. Во время физической работы происходят изменения, способствующие более легкой отдаче кислорода тканям. Повышается васкуляризация мышц (количество сосудов в мышцах, по которым может протекать кровь и доставляться кислород), снижается вязкость крови. Тренировки повышают общую массу гемоглобина путем стимуляции эритропоэза (образования эритроцитов), что увеличивает количество кислорода, которое может переноситься кровью.
Список используемой литературы:
1) Ledingham, 1977; Carroll, 2007.
2) Berglund и Hemmingson, 1987.
3) Eaton et al., 1974; Hebbel et al., 1978.
4) Mairbäurl, 2012.
5) Stamler et al., 1997.
6) Gonzalez-Alonso et al., 2002.
7) Weber и Fago, 2004.
 Laughlin et al., 2012.
Laughlin et al., 2012.
9) Mairbäurl, 1994.
10) Böning et al., 1975; Braumann et al., 1982; Mairbäurl et al., 1983; Schmidt et al., 1988.
11) Mairbäurl et al., 1983.
12) Seamen et al., 1980; Rapoport, 1986; Haidas et al., 1971; Mairbäurl et al., 1990.
13) Mairbäurl и Weber, 2012; Berlin et al., 2002.
14) Böning et al., 1975; Braumann et al., 1982; Mairbäurl et al., 1983.
15) Mairbäurl et al., 1983; Dempsey и Wagner, 1999; Hopkins, 2006; Calbet et al., 2008.
16) Berglund и Hemmingson, 1987.
17) Sawka et al., 2000; Schmidt and Prommer, 2010.
18) Broun, 1922; Davies and Brewer, 1935; Ernst, 1987; Sawka et al., 2000.
19) El-Sayed et al., 2005; Böning et al., 2011.
20) Hu и Lin, 2012.
21) Costill et al., 1974.
22) Sawka et al., 2000.
23) Milledge et al., 1982; Hagberg et al., 1998; Sawka et al., 2000; Heinicke et al., 2001.
24) Sawka et al., 2000.
25) Thirup, 2003.
26) Broun, 1922; Kurz, 1948; Martin и Kilian, 1959.
27) Yoshimura et al., 1980; Miller et al., 1988; Telford et al., 2003; Dressendorfer et al., 1992.
28) Yoshimura et al., 1980; Hunding et al., 1981.
29) Sawka et al., 2000.
30) Kjellberg et al., 1949; Sawka et al., 2000.
31) Parisotto et al., 2000; Schmidt и Prommer, 2008.
32) Heinicke et al., 2001.
33) Kjellberg et al., 1949.
34) Sawka et al., 2000; Schmidt и Prommer, 2008.
35) Hurtado, 1964; Sanchez et al., 1970.
36) Reynafarje et al., 1959; Myhre et al., 1970.
37) Sawka et al., 2000.
38) Schmidt et al., 1988; Convertino, 1991.
39) Lombardi et al., 2013; Banfi et al., 2011; Diaz et al., 2011.
40) Banfi et al., 2011.
41) Shahidi, 1973; Shahani et al., 2009.
42) Hackney, 2001; Enea et al., 2009.
43) Shahani et al., 2009.
44) Dar et al., 2011; Hu and Lin, 2012.
45) Kurtz et al., 1988; Christ et al., 1997.
46) Hakkinen и Pakarinen, 1995; Schwarz et al., 1996.
47) El-Sayed et al., 2005.
48) El-Sayed et al., 2005.
49) Vandewalle et al., 1988; Geor et al., 1994; Yalcin et al., 2000.
50) Van der Brug et al., 1995; Bouix et al., 1998; Smith et al., 2013.
51) Neuhaus et al., 1992.
52) Gurcan et al., 1998.
53) Simmonds et al., 2013.
54) Connes et al., 2004.
55) Romain et al., 2011.
56) Kamada et al., 1993.
57) Mairbäurl et al., 1983; Linderkamp et al., 1993; Pichon et al., 2013; Zhao et al., 2013; Mohandas and Chasis, 1993.
58) Grau et al., 2013.
59) Laughlin et al., 2012.
Во время тренировок потребление кислорода возрастает и чтобы адаптироваться к ним, нам нужно активно доставлять кислород в мышцы. В этом нам помогает гемоглобин. Разберемся, как это работает.
Это часть интерактивных уроков, подготовленных образовательной платформой Level One в сотрудничестве
с крупнейшими российскими экспертами.
Еще 500 уроков по 15 направлениям, от истории
и архитектуры до здоровья и кулинарии на levelvan.ru/plus
посмотреть все уроки

Автор урока
Софья Хабибова
Спортивный травматолог, тренер по фитнесу и нутрициолог, тренер проекта Norma Fit
Перейдем к следующему важному элементу в нашем организме — кислороду. Мы дышим постоянно и жить не можем без кислорода.
Зачем он нам 👇
🦖 Вершина эволюции. Кислород — такой же элемент эволюции, как и жир. Простейшие существа умели без него обходиться, но потом появились растения, которые создавали кислород с помощью солнечной энергии. Благодаря им возникли и более сложные организмы. Сейчас все процессы внутри нас проходят с участием кислорода.
💪 Много энергии. Для работы мышц также нужен кислород. Когда в мышцу приходит жир или белок, мы расщепляем его с помощью кислорода. В процессе расщепления получаем АТФ и тепло. Такие реакции называются аэробными и это самые эффективные процессы — мы получаем максимум АТФ.
🩺 Профилактика болезней. Кислород попадает в мышцы через сердце. Чем больше интенсивность тренировки, тем активнее наше сердце качает кровь. При регулярных тренировках вдох становится глубже, объем легких увеличивается, а в работающих мышцах появляются новые сосуды. Глубокое дыхание помогает нам бороться с сердечно-сосудистыми заболеваниями.
Люди уже давно пытаются понять, насколько эффективно мы тренируемся. Сейчас у многих есть браслет с функцией измерения пульса и этого хватает.
Но есть и другие варианты 👇
🥵 Калориметрия. Сначала эффективность наших тренировок пытались измерить с помощью тепла, которое выделяется при получении энергии. Такой способ называется калориметрия и технически он оказался очень сложным. Тогда ученые переключились на кислород — ведь чем глубже и чаще мы дышим, тем активнее происходят процессы внутри нас.
😮💨 Непрямая калориметрия. Сегодня активность тренировки измеряют с помощью непрямой калориметрии: человека сажают на велосипед или другой кардиотренажер и надевают кислородную маску. Нагрузку увеличивают, пока человек не устанет. Чем тяжелее — тем больше мы вдыхаем. Так можно определить максимум нагрузок, которые может выдержать спортсмен. Калориметрия сегодня почти не используется, а непрямая калориметрия — профессиональный уровень, который нужен не всем.
💗 Пульс. Для повседневных тренировок достаточно следить за частотой пульса. Чем больше нагрузка — тем активнее сердце качает кровь. Пульс — это количество сокращений в минуту. Поэтому когда мы говорим об интенсивности тренировок, мы говорим о частоте сердечных сокращений. Со временем сердце адаптируется к нагрузке и пульс выравнивается — на нагрузках, которые раньше требовали больших значений пульса, мы начинаем показывать меньшие значения пульса.
Если вы хотите активно заниматься спортом, нужно следить за уровнем гемоглобина.
Как это работает 👇
🩸 Как кислород попадает в мышцы. За перенос кислорода и доставку его в клетки отвечает белок гемоглобин. Он содержится в эритроцитах — красных кровяных клетках. Мышцы тоже стараются держать небольшой запас кислорода на всякий случай: для этого есть белок миоглобин. Чем больше у нас гемоглобина, тем проще кислород будет доходить до мышц во время тренировки. Нужно следить за его уровнем и стараться приблизиться к верхней норме.
👨⚕️ Пониженный уровень гемоглобина в организме называют анемией.
Она появляется по разным причинам:
🔹 Дефицит железа в организме;
🔹 Сильная кровопотеря;
🔹 Дефицит витамина B12.
При анемии:
🔹 Человек испытывает слабость;
🔹 Слышит звон в ушах;
🔹 При резком подъеме в глазах темнеет;
🔹 Кожа чаще всего бледная.
❓ Как повысить уровень гемоглобина
🥩 Есть и пить железо. Стоит добавить в свой рацион продукты с высоким содержанием железа: красное мясо, печень и гречку. Также можно пить биологически активные добавки с железом. Железо можно пить без назначения врача. Если у вас низкий гемоглобин или вы активно тренируетесь — смело покупайте железо.
😷 Сократить кислород. Если вы тренируетесь на выносливость и хотите улучшить результаты, то попробуйте сократить количество кислорода во вдыхаемом воздухе во время тренировки. В ответ на такую тренировку организм начинает вырабатывать новые эритроциты и гемоглобин.
Какие есть варианты:
🔸 Тренироваться в душных помещениях. Совет странный, но это работает.
🔸 Надевать на пробежку или тренировку маску. Очень актуально в наше время — и гемоглобин увеличите, и масочный режим не нарушите.
🔸 Заниматься в высокогорье, там где воздух разряжен. Этот вариант доступен не всем, но если поблизости есть горы — советуем выбрать их для пробежек или активной ходьбы.
✋🛑 Допинг — так делать не надо. Последний вариант — неправильный, ведь это допинг. В медицине для роста гемоглобина используют препарат эритропоэтин: им лечат почечную недостаточность. Эритропоэтин стимулирует образование новых красных клеток крови — эритроцитов. Такой препарат надо принимать только по медицинским показаниям, но точно не для того, чтобы сделать тренировки эффективнее.
- Авторы
- Файлы
Чеснокова Н.П.
1
Понукалина Е.В.
1
Моррисон В.В.
1
Бизенкова М.Н.
1
1 ФГБОУ ВО «Саратовский Государственный медицинский университет им. В.И. Разумовского Минздрава России»
4.1. Транспорт кислорода
В сложных механизмах транспорта газов кровью и газообмена в тканях важная роль отводится эритроцитам, ответственным за доставку О2 к различным органам и удаление образующегося в процессе метаболизма СО2.
Эритроцит – безъядерная клетка, лишенная митохондрий, основным источником энергии для эритроцита служит глюкоза, метаболизируемая в гексозомонофосфатном шунте или цикле Эмбдена-Мейергофа. Транспорт О2 обеспечивается в значительной мере гемоглобином, состоящим из белка глобина и гема. Последний представляет собой комплексное соединение железа и порфирина. Глобин представляет собой тетрамер полипептидной цепи. Hb A (HbA) – основной гемоглобин взрослых содержит 2 – альфа и 2 – бета – цепи, Hb A2 – содержит две альфа и две дельта цепи.
Гем состоит из иона железа, встроенного в порфириновое кольцо. Ион железа гема обратимо связывает одну молекулу О2. С одной молекулой Hb максимально связываются 4 молекулы О2 с образованием оксигемоглобина.
Гем может подвергаться не только оксигенации, но и истинному окислению, когда железо становится из двухвалентного трехвалентным. Окисленный гем носит название гематина, а молекула гемоглобина становится метгемоглобином. В крови человека метгемоглобин находится в незначительных количествах, его уровень резко возрастает при отравлениях. Метгемоглобин не способен отдавать кислород тканям.
В норме метгемоглобин составляет менее 3% общего Hb крови. Основная форма транспорта О2 – в виде оксигемоглобина. Кислород транспортируется артериальной кровью не только в связи с гемоглобином, но и в растворенном виде. Принимая во внимание тот факт, что 1 г Hb может связать 1,34 мл О2, кислородная емкость крови в среднем у взрослого человека составляет около 200 мл/л крови. Одним из показателей кислородного транспорта является насыщение артериальной крови О2(Sa O2), равного отношению О2, связанного с Hb, к кислородной емкости крови:
SaO2=O2, связанного с Hb/O2 емкость крови* 100%.
В соответствии с кривой диссоциации оксигемоглобина насыщение артериальной крови кислородом в среднем составляет 97%, в венозной крови – 75%.
PaO2 в артериальной крови около 100 мм. рт. ст., а в венозной – около 40 мм. рт. ст.
Количество растворенного кислорода в крови пропорционально парциальному давлению О2 и коэффициэнту его растворимости.
Последний для О2 составляет 0,0031/100 мл крови/ 1 мм. рт. ст.. Таким образом, 100 мл крови при PaO2, равном 100 мм. рт. ст., содержит менее 0,31 мл O2.
Диссоциация оксигемоглобина в тканях обусловлена главным образом химическими свойствами гемоглобина, а также рядом других факторов – температурой тела, рН среды, р СО2.
При понижении температуры тела наклон кривой диссоциации оксигемоглобина возрастает, а при ее повышении – снижается, и соответственно снижается сродство Hb к О2.
При снижении рН, т.е. при закислении среды, сродство гемоглобина к О2 уменьшается. Увеличение напряжения в крови СО2 также сопровождается снижением сродства Hb к О2 и уплощением кривой диссоциации оксигемоглобина.
Известно, что степень диссоциации оксигемоглобина определяется содержанием в эритроцитах некоторых фосфорорганических соединений, главным из которых является 2,3 – ДФГ (2,3 дифосфоглицерат), а также содержанием в эритроцитах катионов. В случаях развития алкалозов, поглощение О2 в легких увеличивается, но в то же время затрудняется отдача кислорода тканями. При ацидозах наблюдается обратная картина.
4.2.Утилизация кислорода тканями
Тканевое или клеточное дыхание включает три стадии. На первой стадии пируват, аминокислоты и жирные кислоты окисляются до двухуглеродных фрагментов ацетильных групп, входящих в состав ацетилкофермента А. Последние на втором этапе окисления включаются в цикл лимонной кислоты, где происходит образование высокоэнергетических атомов водорода и высвобождение СО2 – конечного продукта окисления органических субстратов. На третьей стадии клеточного дыхания атомы водорода делятся на протоны (Н+) и «высокоэнергетические» электроны, передающиеся по дыхательной цепи на молекулярный О2 и восстанавливающие его до НО2. Перенос электронов сопряжен с запасом энергии в форме АТФ, т.е. с окислительным фосфорилированием (рис.6).
Касаясь патогенеза метаболических сдвигов, свойственных гипоксическим состояниям, следует отметить, что в организме человека более 90% всего потребляемого кислорода восстанавливается с участием цитохромоксидазы митохондрий, и лишь около 10% кислорода метаболизируется в тканях с участием оксигеназ: диоксигеназы и монооксигеназы.
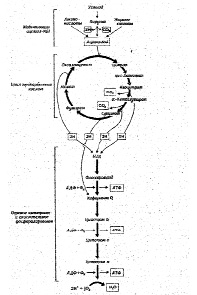
Рис.6. Схема тканевого дыхания. Конечные продукты каждой стадии даны в рамке (Ленинджер А., 1999)
Наиболее многочисленны и сложны монооксигеназные реакции, протекающие в эндоплазматическом ретикулуме клеток при участии цитохрома Р-450 и обеспечивающие гидроксилирование субстрата (стероидных гормонов, лекарственных препаратов и различных др. соединений) и, как правило, его инактивацию.
Диоксигеназы катализируют реакции, в которых в молекулу органического субстрата включаются оба атома молекулы кислорода (например, реакция окисления катехола молекулярным кислородом с раскрытием кольца).
В реакциях, связанных с переносом электронов, т.е. в реакциях окисления-восстановления, где, как указывалось выше, используется более 90% потребляемого кислорода, атомы водорода, отщепленные дегидрогеназами от субстратов в цикле лимонной кислоты, передают свои электроны в цепь переноса электронов и превращаются также в Н +. Как известно, помимо 4 пар атомов водорода, поставляемых каждым оборотом цикла лимонной кислоты, образуются и другие атомы водорода, отщепленные дегидрогеназами от пирувата, жирных кислот и аминокислот в процессе их расщепления до Ацетил-СоА и других продуктов.
Таким образом, все атомы водорода, отщепляемые дегидрогеназами от субстратов, передают свои электроны в дыхательную цепь к конечному акцептору электронов – кислороду.
Касаясь последовательности транспорта электронов в окислительно-восстановительных реакциях, протекающих на внутренней мембране митохондрий, прежде всего, следует отметить, что от всех НАД – зависимых реакций дегидрирования восстановленные эквиваленты переходят к митохондриальной НАДН – дегидрогеназе, затем через ряд железосерных ферментов передаются на убихинон М цитохрому b. Далее электроны переходят последовательно на цитохромы С1 и С, затем на цитохромы аа 3 (цитохромоксидазу – медьсодержащий фермент). В свою очередь цитохромоксидаза передает электроны на кислород. Для того, чтобы полностью восстановить кислород с образованием 2-х молекул воды требуется 4 электрона и четыре Н+ .
Скорость утилизации О2 в различных тканях различна. В среднем взрослый человек потребляет 250 мл О2 в 1 мин. Максимальное извлечение О2 из притекающей артериальной крови свойственно миокарду.
Кислород используется в клетках, в основном в метаболизме белков, жиров, углеводов, ксенобиотиков, в окислительно-восстановительных реакциях в различных субклеточных фракциях: в митохондриях, в эндоплазматическом ретикулуме, в реакциях липопероксидации, а также в межклеточном матриксе и в биологических жидкостях.
Коэффициент утилизации О2 в тканях равен отношению потребления О2 к интенсивности его доставки, широко варьирует в различных органах и тканях.
В условиях нормы минимальную потребность в О2 проявляют почки и селезенка, а максимальную потребность – кора головного мозга, миокард и скелетные мышцы, где коэффициент утилизации О2 колеблется от 0,4 до 0,6, а в миокарде до 0,7. При крайне интенсивной физической работе коэффициент утилизации О2 мышцами и миокардом может возрастать до 0,9.
Обмен дыхательных газов в тканях происходит в процессе свободной и облегченной диффузии. При этом О2 переносится по градиенту напряжения газа из эритроцитов и плазмы крови в окружающие ткани.
Одновременно происходит диффузия СО2 из тканей в кровь. На выход О2 из крови в ткани влияет диссоциация оксигемоглобина в эритроцитах, что обеспечивает так называемую облегченную диффузию О2. Интенсивность диффузионного потока О2 и СО2 определяется градиентом их напряжения между кровью и тканями, а также площадью газообмена, плотностью капилляров, распределением кровотока в микроциркуляторном русле. Интенсивность окислительных процессов в тканях определяется величиной критического напряжения О2 в митохондриях, которое в условиях нормы должно превосходить 0,1-1 мм рт. ст.
Соответствие доставки О2 к органам и тканям, возросшим потребностям в оксигенации обеспечивается на клеточном, органном уровнях за счет образования метаболитов изнашивания, а также при участии нервных, гормональных и гуморальных влияний.
4.3.Транспорт СО2
Основная масса углекислого газа (СО2) образуется в организме как конечный продукт различных метаболических реакций и транспортируется к легким с кровью. Вдыхаемый воздух содержит лишь незначительное количество СО2.
Транспорт СО2 кровью осуществляется в 3-х состояниях: в виде аниона бикарбоната, в растворенной форме и в виде карбаминовых соединений.
СО2 хорошо растворяется в плазме крови и в артериальной крови, около 5% от общей двуокиси углерода содержится в крови в растворенной форме.
Анион бикарбоната составляет около 90% от общего содержания СО2 в артериальной крови: СО2+Н2О — Н++НСО-3.
Эта реакция медленно протекает в плазме крови, но чрезвычайно интенсивно происходит в эритроцитах при участии фермента карбоангидразы. Мембрана эритроцита относительно непроницаема для Н+, как и вообще для катионов, но в тоже время, проницаема для ионов НСО-3, выход которых из эритроцитов в плазму обеспечивается притоком Cl- из плазмы в эритроциты. Часть Н+ забуферивается гемоглобином с образованием восстановленного гемоглобина.
Третьей формой транспорта СО2 кровью являются карбаминовые соединения, образованные взаимодействием СО2 с концевыми группами белков крови преимущественно с гемоглобином:
Hb NH2 + CO2 — Hb NH COOH > Hb NH COO + Н+
Карбаминовые соединения составляют около 5% от общего количества СО2, транспортируемого кровью.
В оксигенированной артериальной крови напряжение СО2 составляет 40 мм. рт. ст., а в венозной крови Рv СО2 равно 46 мм. рт. ст.
4.4.Связывание гемоглобина с окисью углерода
Угарный газ (СО) – окись углерода обладает значительно большим сродством к гемоглобину, чем О2, с последующим образованием карбоксигемоглобина. СО входит в состав бытового газа, а также выделяется при работе двигателей внутреннего сгорания. При концентрации СО во вдыхаемом воздухе всего в количестве 7*10- 4 около 50% гемоглобина превращается в карбоксигемоглобин. В норме в крови содержится около 1% HbCO, у курильщиков – 3% . В крови водителей такси концентрация карбоксигемоглобина достигает 20%. Карбоксигемоглобин диссоциирует с отдачей О2 в 200 раз медленней оксигемоглобина и в то же время препятствует его диссоциации в тканях.
Библиографическая ссылка
Чеснокова Н.П., Понукалина Е.В., Моррисон В.В., Бизенкова М.Н. ЛЕКЦИЯ 4 ФИЗИОЛОГИЯ ТРАНСПОРТА ГАЗОВ КРОВЬЮ И КИСЛОРОДНОГО ОБЕСПЕЧЕНИЯ ТКАНЕЙ // Научное обозрение. Медицинские науки. – 2017. – № 2.
– С. 40-42;
URL: https://science-medicine.ru/ru/article/view?id=973 (дата обращения: 22.03.2023).
Предлагаем вашему вниманию журналы, издающиеся в издательстве «Академия Естествознания»
(Высокий импакт-фактор РИНЦ, тематика журналов охватывает все научные направления)